Download
1 / 2
20 likes | 129 Views
An ERP Investigation of Fast Phonological Priming Kristi Kiyonaga 1 , Jonathan Grainger 2, Maya Misra 3 & Phillip J. Holcomb 1 Tufts University, Medford, MA 1 ; CNRS, Aix/Marseille 2 ; Penn State University 3. Experiment 1. Introduction

E N D
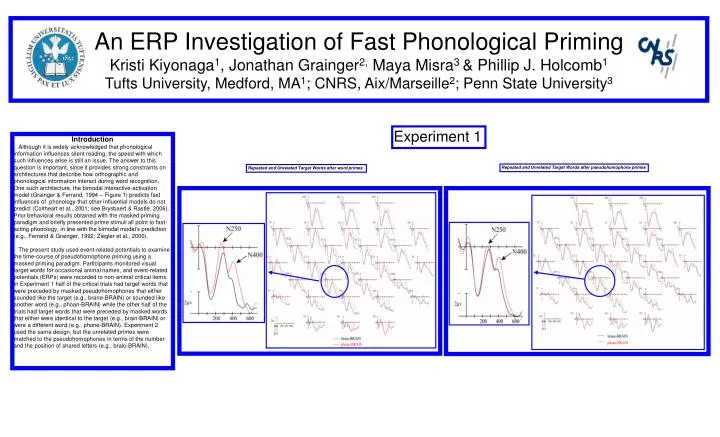
An ERP Investigation of Fast Phonological PrimingKristi Kiyonaga1, Jonathan Grainger2, Maya Misra3 & Phillip J. Holcomb1Tufts University, Medford, MA1; CNRS, Aix/Marseille2; Penn State University3 Experiment 1 Introduction Although it is widely acknowledged that phonological information influences silent reading, the speed with which such influences arise is still an issue. The answer to this question is important, since it provides strong constraints on architectures that describe how orthographic and phonological information interact during word recognition. One such architecture, the bimodal interactive-activation model (Grainger & Ferrand, 1994 – Figure 1) predicts fast influences of phonology that other influential models do not predict (Coltheart et al., 2001; see Brysbaert & Rastle, 2006). Prior behavioral results obtained with the masked priming paradigm and briefly presented prime stimuli all point to fast-acting phonology, in line with the bimodal model’s prediction (e.g., Ferrand & Grainger, 1992; Ziegler et al., 2000). The present study used event-related potentials to examine the time-course of pseudohomophone priming using a masked priming paradigm. Participants monitored visual target words for occasional animal names, and event-related potentials (ERPs) were recorded to non-animal critical items. In Experiment 1 half of the critical trials had target words that were preceded by masked pseudohomophones that either sounded like the target (e.g., brane-BRAIN) or sounded like another word (e.g., phoan-BRAIN) while the other half of the trials had target words that were preceded by masked words that either were identical to the target (e.g., brain-BRAIN) or were a different word (e.g., phone-BRAIN). Experiment 2 used the same design, but the unrelated primes were matched to the pseudohomophones in terms of the number and the position of shared letters (e.g., bralo-BRAIN). Repeated and Unrelated Target Words after pseudohomophone primes Repeated and Unrelated Target Words after word primes
Findings Target words following word (Exp 1) and pseudo-homophone primes (Exp 1 & 2) resulted in significantly smaller amplitudes compared with targets following control primes in two negative-going ERP components peaking at approximately 250 ms post-target onset (N250) and 400 ms post-target onset (N400). That this pattern emerged even when the primes were carefully matched on orthographic features (Exp 2) provides one of the most convincing demonstrations to date of the importance of pre-lexical phonology during silent reading, and helps resolve the long-standing debate on this critical issue. These results are in line with the predictions of the bimodal interactive-activation model (Grainger & Ferrand, 1994 – Figure 1), as well as the recent attempt to map ERP components onto the different component processes in this model (Holcomb & Grainger, in press; Kiyonaga et al., in press). In this scheme the N250 is hypothesized to reflect sub-lexical processing (both orthographic and phonological) whereas the N400 reflects the mapping of whole-word form representations onto their respective meanings. Pseudo-homophone primes affect both of these processes since they activate sub-lexical and whole-word phonological representations that are compatible with upcoming target words. • Method • Experiment 1 - 20 right handed native English speaking participants, 32 Channel EEG (Figure 2) • 260 trials of a semantic categorization task (see Figure 3 top) • 200 critical trials • 60 probe trials – each requiring a button press • 2 critical conditions each with 2 trial types: repeated and unrepeated • Repeated and unrelated target word following a word prime (e.g., brain-BRAIN & phone-BRAIN) • Pseudo-repeated and unrelated target word following a pseudohomophone prime (e.g., brane-BRAIN & phoan-BRAIN) • Experiment 2 - 24 new right handed native English speaking participants • 260 trials of a semantic categorization task (see Figure 3 bottom) • 100 critical trials • 100 filler trials • 60 probe trials – each requiring a button press • 1 critical condition with 2 trial types: pseudo repeated, (brane-BRAIN), and ortho-matchedunrelated (e.g., bralo-BRAIN) • Note that ortho-matched unrelated prime matched with pseudo repeated prime on number and position of shared letters with the target Figure 1 -BIAM Experiment 2 Repeated and Unrelated Target Words after pseudohomophone primes Figure 3 - Trial Schematics • References • Coltheart, M., Rastle, K., Perry, C., Langdon, R., & Ziegler, J. C. (2001). DRC: A dual route cascaded model of visual word recognition and reading aloud. Psychological Review, 108, 204-256. • Ferrand, L. & Grainger, J. (1992). Phonology and orthography in visual word recognition: Evidence from masked nonword priming. Quarterly Journal of Experimental Psychology, 45A, 353-372. • Grainger, J., & Ferrand, L. (1994). Phonology and orthography in visual word recognition: Effects of masked homophone primes. Journal of Memory & Language, 33, 218-233. • Holcomb, P.J. & Grainger, J. (2006). On the time-course of visual word recognition: En ERP investigation using masked repetition priming. Journal of Cognitive Neuroscience, in press. • Kiyonaga, K., Grainger, J., Midgley, K.J., & Holcomb, P.J. (2006). Masked cross-modal repetition priming: An event-related potential investigation. Language and Cognitive Processes, in press. • Rastle, K., & Brysbaert, M. (2006). Masked Phonological Priming Effects in English: Are they Real? Do they Matter? Cognitive Psychology, in press. • Ziegler, J. C., Ferrand, L., Jacobs, A. M., Rey, A., & Grainger, J. (2000). Visual and phonological codes in letter and word recognition: Evidence from incremental priming. Quarterly Journal of Experimental Psychology: Human Experimental Psychology, 53A, 671-692. Figure 2 – Electrode Montage Exp 1 Exp 2 This research was supported by: HD25889 HD043251 & the CNRS






