Download
1 / 1
20 likes | 232 Views
Functions of Acyl-CoA Binding Proteins in C. elegans. Ida Coordt Elle 1 , Karina Trankjær Simonsen 2 , Pernille Kirstine Birck 1 , Thuc Timothy Le 3 , Nils Joakim Færgeman 1. 1 Department of Biochemistry and Molecular Biology, University of Southern Denmark, Odense, Denmark

E N D
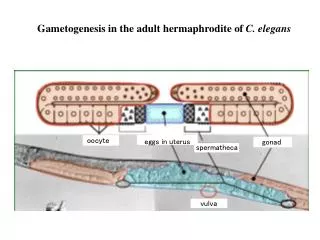
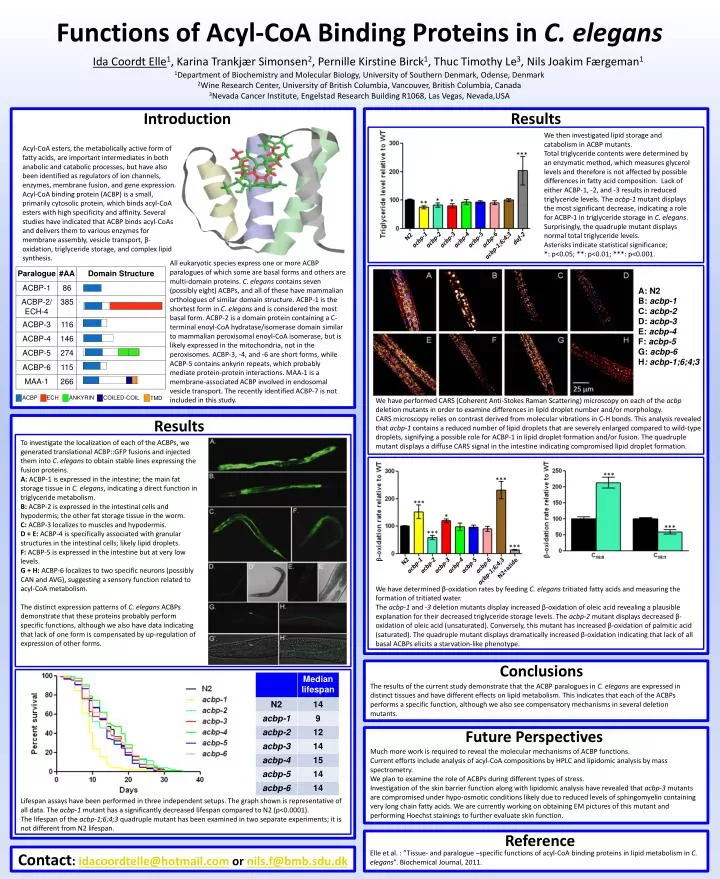
Functions of Acyl-CoA Binding Proteins in C. elegans Ida Coordt Elle1, Karina Trankjær Simonsen2, Pernille Kirstine Birck1, Thuc Timothy Le3, Nils Joakim Færgeman1 1Department of Biochemistry and Molecular Biology, University of Southern Denmark, Odense, Denmark 2Wine Research Center, University of British Columbia, Vancouver, British Columbia, Canada 3Nevada Cancer Institute, EngelstadResearchBuilding R1068, Las Vegas, Nevada,USA Results Introduction Wetheninvestigatedlipidstorage and catabolism in ACBP mutants. Total triglyceride contents were determined by an enzymatic method, which measures glycerol levels and therefore is not affected by possible differences in fatty acid composition. Lack of either ACBP-1, -2, and -3 results in reduced triglyceride levels. The acbp-1 mutant displays the most significant decrease, indicating a role for ACBP-1 in triglyceride storage in C. elegans. Surprisingly, the quadruple mutant displays normal total triglyceride levels. Asterisks indicatestatisticalsignificance; *: p<0.05; **: p<0.01; ***: p<0.001. Acyl-CoA esters, the metabolically active form of fatty acids, are important intermediates in both anabolic and catabolic processes, but have also been identified as regulators of ion channels, enzymes, membrane fusion, and gene expression. Acyl-CoA binding protein (ACBP) is a small, primarily cytosolic protein, which binds acyl-CoA esters with high specificity and affinity. Several studies have indicated that ACBP binds acyl-CoAs and delivers them to various enzymes for membrane assembly, vesicle transport, β-oxidation, triglyceride storage, and complex lipid synthesis. *** * * ** N2 daf-2 acbp-1 acbp-2 acbp-3 acbp-4 acbp-5 acbp-6 acbp-1;6;4;3 All eukaryotic species express one or more ACBP paralogues of which some are basal forms and others are multi-domain proteins. C. eleganscontains seven (possibly eight) ACBPs, and all of these have mammalian orthologues of similar domain structure. ACBP-1 is the shortest form in C. elegans and is considered the most basal form. ACBP-2 is a domain protein containing a C-terminal enoyl-CoAhydratase/isomerase domain similar to mammalian peroxisomalenoyl-CoAisomerase, but is likely expressed in the mitochondria, not in the peroxisomes. ACBP-3, -4, and -6 are short forms, while ACBP-5 contains ankyrin repeats, which probably mediate protein-protein interactions. MAA-1 is a membrane-associated ACBP involved in endosomal vesicle transport. The recently identified ACBP-7 is not included in this study. A: N2 B: acbp-1 C: acbp-2 D: acbp-3 E: acbp-4 F: acbp-5 G: acbp-6 H: acbp-1;6;4;3 ACBP ECH ANKYRIN COILED-COIL TMD We have performed CARS (Coherent Anti-Stokes Raman Scattering) microscopy on each of the acbp deletion mutants in order to examine differences in lipid droplet number and/or morphology. CARS microscopy relies on contrast derived from molecular vibrations in C-H bonds. This analysis revealed that acbp-1 contains a reduced number of lipid droplets that are severely enlarged compared to wild-type droplets, signifying a possible role for ACBP-1 in lipid droplet formation and/or fusion. The quadruple mutant displays a diffuse CARS signal in the intestine indicating compromised lipid droplet formation. Results To investigate the localization of each of the ACBPs, we generated translational ACBP::GFP fusions and injected them into C. elegans to obtain stable lines expressing the fusion proteins. A: ACBP-1 is expressed in the intestine; the main fat storage tissue in C. elegans, indicating a direct function in triglyceride metabolism. B: ACBP-2 is expressed in the intestinal cells and hypodermis; the other fat storage tissue in the worm. C: ACBP-3 localizes to muscles and hypodermis. D + E: ACBP-4 is specifically associated with granular structures in the intestinal cells; likely lipid droplets. F: ACBP-5 is expressed in the intestine but at very low levels. G + H: ACBP-6 localizes to two specific neurons (possibly CAN and AVG), suggesting a sensory function related to acyl-CoA metabolism. The distinct expression patterns of C. elegansACBPs demonstrate that these proteins probably perform specific functions, although we also have data indicating that lack of one form is compensated by up-regulation of expression of other forms. *** *** *** * *** *** *** N2 acbp-1 acbp-2 acbp-3 acbp-4 acbp-5 acbp-6 N2+azide acbp-1;6;4;3 We have determined β-oxidation rates by feeding C. eleganstritiated fatty acids and measuring the formation of tritiated water. The acbp-1 and -3 deletion mutants display increasedβ-oxidation of oleicacidrevealing a plausible explanation for theirdecreasedtriglyceridestoragelevels. The acbp-2 mutant displays decreased β-oxidation of oleic acid (unsaturated). Conversely, this mutant has increasedβ-oxidation of palmiticacid (saturated).The quadruple mutant displays dramaticallyincreasedβ-oxidation indicatingthatlack of all basal ACBPs elicits a starvation-like phenotype. Conclusions The results of the current study demonstrate that the ACBP paralogues in C. elegansare expressed in distinct tissues and have different effects on lipid metabolism. This indicates that each of the ACBPs performs a specific function, although we also see compensatory mechanisms in several deletion mutants. Future Perspectives Much more work is required to reveal the molecular mechanisms of ACBP functions. Currenteffortsincludeanalysis of acyl-CoAcompositions by HPLC and lipidomicanalysis by massspectrometry. We plan to examine the role of ACBPs duringdifferent types of stress. Investigation of the skin barrierfunctionalongwithlipidomicanalysis have revealedthatacbp-3 mutants arecompromised under hypo-osmoticconditionslikely due to reducedlevels of sphingomyelincontainingvery long chainfattyacids. Wearecurrentlyworkingonobtaining EM pictures of this mutant and performing Hoechst stainings to furtherevaluate skin function. Lifespan assays have been performed in three independent setups. The graph shown is representative of all data. The acbp-1 mutant has a significantly decreased lifespan compared to N2 (p<0.0001). The lifespan of the acbp-1;6;4;3 quadruple mutant has been examined in two separate experiments; it is not different from N2 lifespan. Reference Elle et al. : ”Tissue- and paralogue –specificfunctions of acyl-CoA binding proteins in lipidmetabolism in C. elegans”. Biochemical Journal, 2011. Contact: idacoordtelle@hotmail.comornils.f@bmb.sdu.dk