Download

1 / 33
340 likes | 598 Views
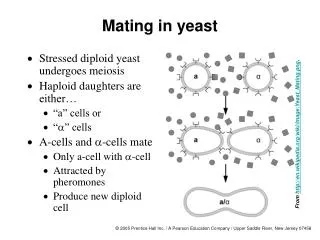
UV Mutagenesis in Yeast. Geneticists need variation to study the function of gene products. We create variation in the laboratory by mutagenesis. Ease of Genetic Manipulation. proteins. 4. 3. 2. 6. 5. DNA. 1. Features of a good model organism. Cell biology.

E N D
UV Mutagenesis in Yeast Geneticists need variation to study the function of gene products. We create variation in the laboratory by mutagenesis
Ease of Genetic Manipulation proteins 4 3 2 6 5 DNA 1 Features of a good model organism • Cell biology • Conserved cellularprocesses • Biochemistry
Important lab properties • Genetics: introduce mutations (UV, chemical, Xray) and design screens to identify mutations in process you are interested in • Isolate the products of meiosis • Recover mutations: stable haploid and diploid lifecycles. • Easy molecular manipulations-clonability (few introns), high rate of homologous recombination
Fig. 7.2 Mutations are classified by what they do to DNA
By choosing the correct mutagens, we can control the type of mutations we make. Because most of the chemicals are nasty, you will be using UV light to generate mutations
sugar sugar sugar Formation of Thymidine Dimer sugar
Photoreactivation requires photolyase enzyme Not present in humans
Mutagenesis of yeast Haploid Irradiate with UV. Calculate survival curve Select optimal dose for isolation of mutations. Select on appropriate selective media: Replica plating to identify nutrient deficiencies.
Survival curve In yeast, 50-90% killing is used in mutagenesis experiments
DNA damage is more lethal to haploids Haploids Diploid cells Multicellular diploids
What can you do with yeast if you are not a geneticist? • Take advantage of the genes identified in genetic screens • Cell biology
Fusions to GFP allow live cell analysis of protein function Aequoria victoria Green Fluorescent Protein
Visualization of chromosome and microtubule dynamics to elucidate mechanisms of nuclear movement and chromosome segregation • lacI-GFP binding to tandem repeat lacO sequences integrated at specific loci. • Dynamics of GFP-fusions to tubulin, spindle pole and mt-based motor proteins. • Specific genetic perturbations to establish cause and effect
Yeast Mitotic Spindle Structure EM Tub-GFP Kubai, 1978 Tomographic reconstruction 16 kinetochore microtubules and 4 interpolar microtubulesemanate from each spindle pole: 40 mts/1.5 m spindle
microtubule Architecture of the kinetochore Maiato et al. (2004) KINETOCHORE CENTROMERE MT PLUS ENDS FIBROUSCORONA OUTER PLATE Ndc80/Hec1 Nuf2 Spc24 Spc25 BUB1 BUBR1 BUB3 MPS1 MAD1 MAD2 Cdc20 CENP-E KinI,Kip3? CENTROMERIC HETEROCHROMATIN CENP-B MCAK INCENP Aurora B Survivin Borealin/Dasra B ICIS INNER PLATE INTERZONE CENP-A CENP-C CENP-G CENP-H CENP-I/hMis6 CeKNL-1 hMis12 3F3/2 antigens McEwen et al. (1998) tension receptors & checkpoint signalling chromatid pairing, structural support, & MT attachment error correction kinetochore assembly & size determination MT attachment, regulation of MT dynamics, & checkpoint signalling OUTER KINETOCHORE INNER KINETOCHORE
Ndc80RFP Spc29RFP overlay Smc3GFP DIC Ndc80RFP overlay Smc3GFP DIC Kinetochores are clustered at the ends the cohesin cylinder
Interstrand cohesin Inflection point Kinetochore sleeve Intrastrand cohesin C-loop Proposed Path of Centromere DNA in a Eukaryotic Kinetochore: C-loop
Patterns observed at 37oC and 25oC in temperature sensitive mutants
Effects of Centromeric Attachment: ndc10-1 at Restrictive Temperature Centromere Microtubule
Effects of Microtubule Attachment: nuf2-60 at Restrictive Temperature Centromere Microtubule
Effects of Microtubule Attachment on the Inner and Outer Kinetochore Restrictive Temperature Permissive Temperature
Using genetics and cell biology we have dissected functional elements of the kinetochore Inner kinetochore complex CBF3 bends centromere DNA . Clustering of 16 kinetochores requires both inner (COMA, MIND) and outer kinetochore (Ndc80) complexes. Outer kinetochore complexes essential for plus end microtubule interactions. Anderson et al. MCB 2009