Download
1 / 18
190 likes | 351 Views
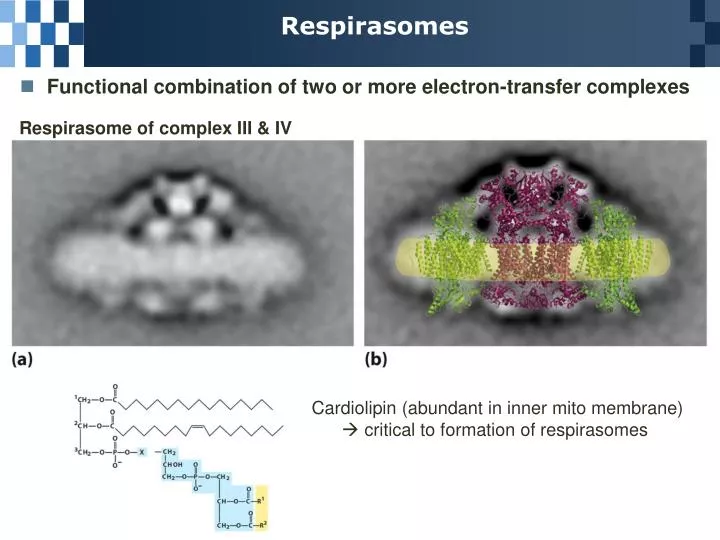
Respirasomes. Functional combination of two or more electron-transfer complexes. Respirasome of complex III & IV. Cardiolipin (abundant in inner mito membrane) critical to formation of respirasomes. Conservation of the e - Transfer Energy in a Proton Gradient.

E N D
Respirasomes • Functional combination of two or more electron-transfer complexes Respirasome of complex III & IV Cardiolipin (abundant in inner mito membrane) critical to formation of respirasomes
Conservation of the e- Transfer Energy in a Proton Gradient • Standard free energy change of e- from NADH to O2 • NADH + H+ + ½ O2 NAD+ + H2O • DG’o = -nFDE’o = -220 kJ/mol • Oxidation of succinate • DG’o = -150 kJ/mol • Using the energy to pump protons out of the matrix • 4H+ (complex I), 4H+ (complex III), 2H+ (complex IV) ; NADH + 11 HN+ + 1/2O2 NAD+ + 10Hp++ H2O
Conservation of the e- Transfer Energy in a Proton Gradient • Electrochemcial energy proton-motive force • Energy stored in proton gradient • Two components • Chemical potential energy : separation of chemical H+ • Electrical potential energy : separation of charge
Proton-Motive Force • Free energy change for the creation of electrochemical gradient by ion pump • DG = RTln(C2 / C1) + ZFDy • C : concentrations of an ion, C2 > C1 • Z : absolute value of electrical charge (1 for a proton) • Dy : transmembrane difference in electrical potential • ln(C2 / C1) = 2.3 (log [H+]P - log [H+]N) = 2.3 (pHN –pHP) = 2.3 DpH • DG = 2.3 RTDpH +FDy • Active mitochondria ; Dy = 0.15 ~ 0.2 V, DpH = 0.74 DG = 19 kJ/mol (of H+) • ~ 200 kJ/mol (NADH)
Reactive oxygen species (ROS) • ROS generation of oxidative phosphorylation • Radical •Q- intermediate • generated during complex I QH2 • QH2 complex III • Pass an electron to O2 • O2 + e- •O2- (superoxide) • Detoxification systems • Superoxide dismutase (SOD) • Glutathione peroxidase (GPx)
Oxidation of NADH in plant mitochondria • Analogous to mito ATP synthesis mechanism of animal • Plant specific alternative mechanism • Regeneration of NAD+ from unneeded NADH • Direct e- transfer from ubiquinone to O2 (bypassing complex III and complex IV) • Energy from e- transfer heat generation • CN- alternative QH2 oxidase
Oxidative Phosphorylation 19.2 ATP Synthesis
Chemiosmotic Model “ The electrochemical energy inherent in the difference in proton concentration and the separation of chargeacross the inner mitochondrial membrane – the proton-motive force– drives the synthesis of ATP as protons flow passively back into the matrix through a proton pore associated with ATP synthase” Chemical potential ∆ pH Electrical potential ∆ ψ
Coupling of Electron Transfer & ATP Synthesis • Experiment to demonstrate ‘coupling’ ADP + Pi succinate • Substrate oxidation • O2 consumption • ATP synthesis Isolated mitochondria Experiment 1 • (ADP + Pi) addition no respiration & ATP synthesis • Succinate addition respiration & ATP synthesis • CN- addition inhibiting respiration & ATP synthesis • Coupling of electron transfer & ATP synthesis
Coupling of Electron Transfer & ATP Synthesis Experiment 2 • Succinate addition no respiration & ATP synthesis • (ADP + Pi) addition respiration & ATP synthesis • Oligomycin or venturicidin addition inhibiting respiration & ATP synthesis • (4) DNP addition continuing respiration without ATP synthesis (uncoupling) • Further demonstration of coupling of electron transfer & ATP synthesis
Coupling of Electron Transfer & ATP Synthesis • Chemical uncouplers • 2,4-dinitrophenol (DNP) & carbonylcyanide-ρ-trifluoromethoxyphenylhydrazone (FCCP) • Weak acids with hydrophobic properties • Release protons in the matrix dissipation of proton gradient • Ionophores • Valinomycin (peptide ionophore binding K+) • Transport of inorganic ions through membranes dissipation of electrical gradient
Evidence for the Role of a Proton Gradient in ATP Synthesis • Artificial generation of electrochemical gradient Leads to ATP synthesis without oxidizable substrate
Mechanism of ATP formation 1. Karl Lohman (1929) 2. Fritz Lipmann (1953) 3. Efraim Racker (1960) 4. Peter Mitchell (1961) 5. Masasuke Yoshida (1997) 6. Paul D. Boyer How is the pmf transmitted to the ATP synthesis? FoF1-ATPase
ATP Synthase • Mitochondrial ATP synthase (complex V) • F-type ATP synthase • Similar to ATP synthase of chloroplast and bacteria • Two functional domains • F1 : peripheral membrane protein • ATP synthesis • Isolated F1 : ATP hydrolysis (originally called F1 ATPase) • Fo (o : oligomycin-sensitive) • Membrane integrated • Proton pore
ATP Synthase : F1 • A3b3gde (9 subunits) • Knoblike structure with alternating a and b arrangement • b subunits; catalytic sites for ATP synthesis • g subunit • Central shaft • Association with one of the three b subunits (b-empty) • Induction of conformational difference in b subunits difference in ADP/ATP binding sites of b subunits b-ATP, b-ADP, b-empty
ATP Synthase : Fo • ab2c10-12 (3 subunits) • C subunit • Small (Mr 8,000), hydrophobic two transmembrane helices • Two concentric circles • Inner circle : N-terminal helices • Outer circle (55 Å in diameter) : C-terminal helices
Mechanism of ATP synthesis in F1 • 18O-exchange experiment with purified F1 • Incubation of purified F1 with ATP in 18O-labelled water Analysis of 18O incorporation into Pi 3 or 4 isotopes in Pi • Repetitive reaction of both ATP hydrolysis and ATP synthesis ; ADP + Pi ATP + H2O, DG’o ≈ 0 reversible reaction!
Mechanism of ATP synthesis in F1 • Kinetic study for confirmation of DG’o ≈ 0 Enz-ATP Enz-(ADP + Pi) • K’eq = k-1/k1 = 24 s-1/10 s-1 = 2.4 DG’o ≈ 0 differ from ATP (free in solution) hydrolysis DG’o = -30.5 kJ/mol (K’eq=105) • FoF1 has high affinity to ATP (Kd < 10-12 M) than ADP (Kd ≈ 10-5 M) 40 kJ/mol difference in binding energy • Equilibrium toward ATP synthesis • Release of ATP from the enzyme surface ; major energy barrier (not ATP formation) Proton gradient makes it possible
