Download

1 / 32
320 likes | 485 Views
Architectures of Mammalian and Fungal Fatty Acid Synthases Presentation based on: T. Maier, S. Jenni, N. Ban, Science 311 , 1258 (2006). -- Mammalian fatty acid at 4.5 Å resolution S. Jenni, M. Leibundgut, T. Maier, N. Ban, Science 311 , 1263 (2006).

E N D
Architectures of Mammalian and Fungal Fatty Acid Synthases Presentation based on: T. Maier, S. Jenni, N. Ban, Science311, 1258 (2006). -- Mammalian fatty acid at 4.5 Å resolution S. Jenni, M. Leibundgut, T. Maier, N. Ban, Science311, 1263 (2006). -- Fungal fatty acid at 5 Å resolution
Agenda 1. fatty acid quick peak 2. catalytic cycle of fatty acid synthesis 3. mammalian fatty acid synthase structure 4. fungal fatty acid synthase structure
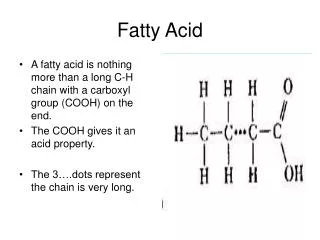
1. Fatty acid quick peak Common fatty acids are carboxylic acids with long hydrocarbon tails: comes with a COOH head and a tail of many CH2.
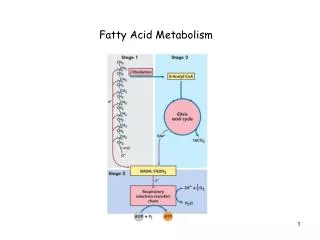
2. Fatty acid catalytic cycle 2.1 Common elongation scheme starter substrates Acetyl coenzyme A (Acetyl-CoA) and Malonyl-CoA transfer the active functionals to acyl carrier protein (ACP). ACP transports substrate to different reaction sites, catalyzed by different enzyme. A complete cycle gives the acyl group an additional two carbon units. This step-wise elongation repeats until a substrate length of C16 to C18 is achieved. Another enzyme then release the substrate from ACP, completing the synthesis process.
2.2 Step-by-step details of catalytic cycle :catalysts for different individual reactions A F B protein that releases the completed product from ACP. C E D
2.3 Different fatty acid synthase (FAS) systems Type II FAS (bacteria) -- all reactions carried out by individual, monofunctional proteins. Type I FAS (eukaryote) -- large, multifuctional polypeptides contains all necessary enzymes for the enlongation cycle. Fungal FAS: 2.6-MD 66 dodecamer, catalytic domains distributed over two distinct subunits. Vertebrates and Mammal FAS: 270-kD2 homodimer, contains all catalytic activities.
3. Mammalian fatty acid synthase 3.1 Functional proteins ACP: acyl protein carrier MAT: malonyl-CoA-/acetyl- CoA-ACP-transacylase KS: -ketoacyl synthase KR: -ketoacyl reductase DH: dehydratase ER: -enoyl reductase TE: thioesterase
3.2 overall structure and domain assignment 210Å 180Å 90Å Blue bubble: Electron density cloud via X-ray crystallography Colored proteins: Identified as specific domain via mapping homologous protein structures
3.2.1 KS domain Mammalian KS closely resembles the Escherichia coli KS I (FabB). So KS domain was fitted with E. coli FabB.
3.2.2 MAT domain Mammalian MAT is homologous to bacterial malonyl transferase (FabD). So we fit the MAT domain with Streptomyces coelicolor FabD.
3.2.3 DH domain Mammalian DH adopts a “double hot dog” fold that’s closely related to the fold of the dimeric bacterial dehydratases FabA and FabZ. So we fit DH with two monomers of dimeric E. coli FabA.
3.2.4 ER domain The best structural match for ER was obtained with a zinc-free bacterial quinone reductase. Here the particular model is quinone reductase of T. thermophilus.
3.2.5 KR domain KR belongs to the short-chain dehydrogenase family, and was modeled with E. coli FabG.
3.2.6 ACP and TE domain * ACP and TE domains could not be placed with confidence, likely due to their inherent flexibility. * However… This blurred volume of electron density, which was observed only on one side, might be interpreted as arising from the C-terminal ACP and TE domains.
3.3 intersubunit and interdomain connections FAS is an intertwined dimer with a large dimerization interface. KS domain dimerize in the same way as homologous homodimeric FabB.
The interaction between ER monomers are guided by the formation of a continuous 12-stranded -sheet. The same way as the homologous bacterial enzyme.
COLORED: identified domains GREY: unassigned region. There are other substantial intersubunit contacts in the unassigned region of electron density map.
3.4 active sites and the two reaction chambers Solid spheres: active sites Hollow spheres: radii = length of the phosphopantheteine arm of ACP
4. Fungal fatty acid synthase 4.1 Functional proteins ACP: acyl protein carrier MPT: malonyl/palmitoyl transferase KS: ketoacyl synthase KR: ketoacyl reductase DH: dehydratase ER: enoyl reductase AT: acetyl transferase PT: phosphopantetheine transferase (for ACP activation)
4.2 overall structure and domain assignment 230Å 230Å 260Å MPT: malonyl/palmitoyl transferase KS: ketoacyl synthase KR: ketoacyl reductase DH: dehydratase ER: enoyl reductase AT: acetyl transferase ACP and PT structures could not be identified. White regions denote unidentified electron density.
4.2.1 KS domain KS dimer domain was identified by finding the thiolase fold in the FAS electron density map (bacterial KS is known to adopt a thiolase fold and form homodimers). Bacterial KS homolog fits almost perfectly into the density map.
4.2.2 KR domain The 4-helix bundle is a characteristic trait of one of the dimerization interface in type-II tetrameric KR homolog of Brassica napus. It also contains a Rossmann fold. The Brassica napus KR homolog fits FAS electron density remarkably well.
4.2.3 DH domain The closest sequence homolog with known structure of fungal DH is the human Peroxisomal 2-enoyl-CoA hydratase 2 involved in oxidation of fatty acids. It is a pseudo-dimer, has two “hot dog” folds, and forms a large sheet. DH structures of bacterial FAS fits less well, since they are true homodimers.
4.2.4 ER domain The fungal ER, unlike other FAS systems, is a FMN-containing oxidoreductase, and no homology has been observed. Since a TIM-barrel fold was discovered near the FMN-binding pocket, the 21 known TIM- barrel superfamilies were examined, and a good fit was obtained with spinach glycolate oxidase.
4.2.5 AT and MPT AT MPT AT and MPT are homologous in sequence, catalyze similar reactions, and have same protein fold. Therefore they are both fitted with malonyl transferase from Streptomyces coelicolor.
In order to unambiguously assign AT and MPT domain, the locations of their N termini relative to the C terminus of DH were observed.
4.3 The reaction chambers COLORED: fitted domains GREY: unassigned region. ** Two identical reaction chambers separated by central wheel. ** Each contains three copies of a full set of catalytic domains. ** All active sites are oriented towards interior. Red cones indicate entrances to active sites.
4.3 The reaction chambers Green sphere: reaction chamber center. A set of active sites in the reaction chamber with all enzymatic activities required for the synthesis cycle.
4.3 The reaction chambers Schematic path of ACP, shuttling substrate between the active sites.