Download

1 / 59
710 likes | 1.08k Views
Oncogenes and tumour suppressor genes. Dr Orla Sheils, Senior Lecturer in Molecular Pathology, Department of Histopathology. Introduction. cell division process is dependent on a tightly controlled sequence of events.

E N D
Oncogenes and tumour suppressor genes Dr Orla Sheils, Senior Lecturer in Molecular Pathology, Department of Histopathology.
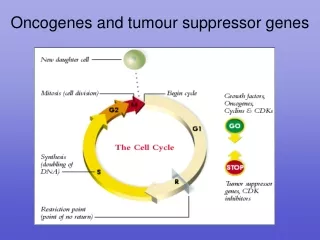
Introduction • cell division process is dependent on a tightly controlled sequence of events. • dependent on the proper levels of transcription and translation of certain genes. • When this process does not occur properly, unregulated cell growth may be the end result.
Introduction • Of the 30,000 or so genes that are currently thought to exist in the human genome, there is a small subset that seems to be particularly important in the prevention, development, and progression of cancer. • These genes have been found to be either malfunctioning or non-functioning in many different kinds of cancer. • The genes that have been identified to date have been categorized into two broad categories, depending on their normal functions in the cell.
Genes whose protein products stimulate or enhance the division and viability of cells. This first category also includes genes that contribute to tumour growth by inhibiting cell death. • Genes whose protein products can directly or indirectly prevent cell division or lead to cell death.
The normal versions of genes in the first group (whose protein products stimulate or enhance the division and viability of cells )are called proto-oncogenes. • The mutated or otherwise damaged versions of these genes are called oncogenes. • The genes in the second group (whose protein products can directly or indirectly prevent cell division or lead to cell death) are called tumour suppressors.
Definition and discovery • Cancer is caused by an accumulation of genetic alterations that confer a survival advantage to the neoplastic cell. • Genetic Changes affect multiple facets: • Cell proliferation • Apoptosis • Tissue invasiveness • Production of growth and angiogenic factors • Ability to escape immune surveillance
Genetic Basis for Cancer • Reflected in the clonal nature of neoplastic cells • Polyclonal growth/ hyperplasia • Response to an • Extrinsic growth factor or • Internal genetic mutation shared by • All cells - MEN2a –germline ret mutation • Some cells – McCune Albright, postzygotic somatic Gsa mutation. • Hyperplastic cells may subsequently acquire one or more somatic mutations and develop clonal derivatives.
Genetic Basis for Cancer • Reflected in the clonal nature of neoplastic cells • Monoclonal growth reflects the acquisition of somatic mutations that confer survival advantage.
tumour suppressors function in many key cellular processes including the regulation of transcription, DNA repair and cell:cell communication. • The loss of function of these genes leads to abnormal cellular behavior.
Cancer – a genetic disease • Intensive effort to characterise genetic alterations associated with different forms of cancer • Landmark advances: • Recognition that some cancers ‘run in families’ • 1900s- Rous sarcoma virus (RSV) • sarcoma transmission in chickens • Retrovirus • Harboured a abnormal variant of a normal cellular gene src • Viral gene product referred to as an oncogene • Greek onkos = mass or tumour • Many viral oncogenes correspond to altered versions of normal cellular genes (proto-oncogenes) [src, ras, raf, kit, jun, fos…].
DNA tumour viruses • Important role in current understanding of neoplasia. • Viruses produce proteins - target key cellular regulatory proteins • Rb, p53 • SV40 large T Ag associates with and inactivates Rb. • Adenovirus E1A also targets Rb
Hereditary Cancers • Transmitted in autosomal dominant manner. • Based on age dependent appearance of retinoblastoma –Knudson postulated a “two-hit” model for the disease. • Hit 1 = germline inherited mutation in Allele 1 • Hit 2 = somatic event involving remaining normal allele.
Tumour Suppressor Genes • Historically –suspected based on several lines of evidence: • Malignant phenotype suppressed by fusion with normal cells (presence of tumour suppressor in normal implied). • Chromosomal losses in hybrids caused reversion to malignant phenotype. • Introduction of single chromosomes into malignant cells: • e.g. insertion of chromosome 11( WT-1 gene) could suppress tumourigenicity in Wilm’s tumour cell line.
tumour Suppressor Genes • Some genes suppress tumour formation. • Their protein product inhibits mitosis. • When mutated, the mutant allele behaves as a recessive; that is, as long as the cell contains one normal allele, tumour suppression continues. • (Oncogenes, by contrast, behave as dominants; one mutant, or overly-active, allele can predispose the cell to tumour formation).
Example 1: RB - the retinoblastoma gene • Retinoblastoma is a cancerous tumour of the retina. It occurs in two forms: • Familial retinoblastoma • Multiple tumours in the retinas of both eyes occurring in the first weeks of infancy. • Sporadic retinoblastoma • A single tumour appears in one eye sometime in early childhood before the retina is fully developed and mitosis in it ceases. • Familial retinoblastoma • Familial retinoblastoma occurs when the fetus inherits from one of its parents a chromosome (number 13) that has its RB locus deleted (or otherwise mutated). The normal Rb protein prevents mitosis.
Mechanism. The Rb protein prevents cells from entering S phase of the cell cycle. It does this by binding to a transcription factor called E2F. • This prevents E2F from binding to the promoters of such proto-oncogenes as c-myc and c-fos. • Transcription of c-myc and c-fos is needed for mitosis so blocking the transcription factor needed to turn on these genes prevents cell division.
Retinoblastoma • A random mutation of the remaining RB locus in any retinal cell completely removes the inhibition provided by the Rb protein, and the affected cell grows into a tumour. So, in this form of the disease, a germline mutation plus a somatic mutation of the second allele leads to the disease.
Example 2: p53 • The product of the tumour suppressor gene p53 is a protein of 53 kilodaltons (hence the name). • The p53 protein prevents a cell from completing the cell cycle if • its DNA is damaged or • the cell has suffered other types of damage. • When • the damage is minor, p53 halts the cell cycle — hence cell division — until the damage is repaired. • the damage is major and cannot be repaired, p53 triggers the cell to commit suicide by apoptosis.
These functions make p53 a key player in protecting us against cancer; that is, an important tumour suppressor gene. • More than half of all human cancers do, in fact, harbour p53 mutations and have no functioning p53 protein.
Loss Of Heterozygosity (LOH) • Because tumour suppressor genes are recessive, cells that contain one normal and one mutated gene — that is, are heterozygous — still behave normally. • However, there are several mechanisms which can cause a cell to lose its normal gene and thus be predisposed to develop into a tumour. These may result in a "loss of heterozygosity" or "LOH".
Mechanisms of LOH: • Deletion of • the normal allele; • the chromosome arm containing the normal allele; • the entire chromosome containing the normal allele (resulting in aneuploidy). • Loss of the chromosome containing the normal allele followed by duplication of the chromosome containing the mutated allele. • Mitotic recombination. The study of tumour suppressor genes revealed (for the first time) that crossing over — with genetic recombination — occasionally occurs in mitosis (as it always does in meiosis). In #2 and #3, the resulting cell now carries two copies of the "bad" gene. This is called "reduction to homozygosity".
Methylation • Mutation is not the only way to inactivate tumour suppressor genes. • Their function can also be blocked by methylation of their promoter.
Mechanism of parental imprinting • The process of imprinting start in the gametes where the allele destined to be inactive in the new embryo (either the father's or the mother's as the case may be) is "marked". The mark appears to be methylation of the DNA in the promoter(s) of the gene. • Methyl groups are added to cytosines (Cs) in the DNA. • occurs at stretches of alternating Cs and Gs called CpG islands. • Methylation of promoters prevents binding of transcription factors to the promoter thus shutting down expression of the gene.
Methylation • Cancer cells often contain a methylated promoter on one tumour suppressor gene accompanied by • a similarly blocked promoter on the other allele (producing the same effect as #2 above); • a loss of that locus on the other chromosome (like the LOH in #1 above); • an inactivating mutation in the other allele.
tumour suppressor genes = anti-oncogenes • Genes like RB and p53 are also called anti-oncogenes. They were first given this name because they reverse, at least in cell culture, the action of known oncogenes.
Tumour suppressor genes • This image (courtesy of Moshe Oren, from Cell 62:671, 1990) shows petri dishes which were seeded with the same number of cells that had been transformed by two oncogenes: myc and ras. • Many of those on the left have grown into colonies of cells. • However, the cells plated on the right also contained the tumour suppressor p53 gene. Only a few have been able to grow into colonies.
Human Papillomavirus (HPV) • The name anti-oncogene may be even more appropriate than originally thought. • Both the Rb protein and the p53 protein complex directly in the cell with an oncogene product of some human papilloma viruses. • Once inside the cells of their host, human papilloma viruses synthesise • a protein designated E7 and • another designated E6.
Human Papillomavirus • The E7 protein of one of these binds to the Rb protein preventing it from binding to the host transcription factor E2F. • Result: E2F is now free to bind to the promoters of genes (like c-myc) that cause the cell to enter the cell cycle . Thus this version of E7 is an oncogene product. • The E6 protein of human papilloma virus implicated in cervical cancer binds the p53 protein targeting it for destruction by proteasomes and thus removing the block on the host cell's entering the cell cycle.
Oncogenes • Genes associated with the stimulation of cell division. • Cancers result from only one mutant allele of gene.
Oncogenes • Growth Factors or Receptors for Growth Factors • PDGF Platelet Derived Growth Factor (brain and breast cancer) • erb-B receptor for epidermal growth factor (brain and breast cancer) • erb-B2 receptor for growth factor (breast, salivary, and ovarian cancers) • RET growth factor receptor (thyroid cancer)
Oncogenes 2. Cytoplasmic Relays in Stimulatory Signaling Pathways • Ki-ras activated by active growth factor receptor proteins (lung, ovarian, colon and pancreatic cancer) • N-ras activated by active growth factor receptor proteins (leukaemias) • c-src is a protein kinase that becomes overactive in phosphorylation of target proteins
Oncogenes 3. Transcription Factors that Activate Growth Promoting Genes • c-myc activates transcription of growth stimulation genes (leukemia, breast, stomach, and lung cancer) • N-myc (nerve and brain cancer) • L-myc (lung cancer) • c-jun and c-fos function as transcription factors
Oncogenes 4. Other types of molecules • Bcl-2 normal protein blocks cell suicide (lymphoma) • Bcl-1 codes for cyclin D1, stimulatory protein of the cell cycle (breast, neck, head cancers) • MDM2 codes for antagonist of p53 (sarcomas)
RAS • Ras gene products are involved in kinase signalling pathways that control the transcription of genes, which then regulate cell growth and differentiation. • To turn "on" the pathway, the ras protein must bind to a particular molecule (GTP) in the cell. • To turn the pathway "off," the ras protein must break up the GTP molecule. • Alterations in the ras gene can change the ras protein so that it is no longer able to break up and release the GTP.
These changes can cause the pathway to be stuck in the "on" position. • The "on" signal leads to cell growth and proliferation. • ras overexpression and amplification can lead to continuous cell proliferation, which is a major step in the development of cancer. • Cell division is regulated by a balance of positive and negative signals. • When ras transcription is increased, an excess of the gene's protein is in the cell, and the positive signals for cell division begin to outweigh the negative signals.
RAS • The conversion of ras from a proto-oncogene into an oncogene usually occurs through a point mutation in the gene. • The altered function can affect the cell in different ways because ras is involved in many signaling pathways that control cell division and cell death. • Anti-cancer drugs are now being developed that target ras dependent pathways. Much remains to be discovered before these drugs can be put into use
RAS • Mutant ras has been identified in cancers of many different origins, including: pancreas (90%), colon (50%), lung (30%), thyroid (50%), bladder (6%), ovarian (15%), breast, skin, liver, kidney, and some leukaemias.
MYC • The myc protein acts as a transcription factor and it controls the expression of several genes. • Mutations in the myc gene have been found in many different cancers, including Burkitt's lymphoma, B-cell leukemia, and lung cancer. • The myc family of oncogenes may become activated by gene rearrangement or amplification.
Gene rearrangements involve the breakage and re-sealing of chromosomes. • This process can involve large amounts of DNA and can affect many genes. • The movement of a gene or group of genes to a different location within the same chromosome or to a different chromosome often leads to altered gene expression and cell function.
SRC • The Src protein is a tyrosine kinase. • Kinases are enzymes that transfer phosphate groups onto target molecules. • The important aspect of this process is that the removal/addition of phosphates changes biomolecules and is a key way by which the activities of cells are regulated.
SRC • The phosphate addition/removal process acts like an on/off switch to control the activity of the target molecules. • The src proteins alter several target molecules, resulting in the transmission of signals to the nucleus that help regulate the cell
Tyrosine Kinases • MAP kinase (MAPK) signaling is among centralsignaling pathways that regulate cell proliferation, celldifferentiation and apoptosis. • As MAPK should transmitextracellular signals to proper regions or compartments incells, controlling subcellular localisation of MAPK is importantfor regulating fidelity and specificity of MAPKsignaling.