Download
1 / 35
410 likes | 761 Views
Bi/CNS 150 Lecture 21 Friday November 15, 2013 Learning & Memory 1. Synaptic plasticity Henry Lester. Chapters 66, 67. The Hebbian Synapse. From The Organization of Behavior by Donald Hebb, 1949:

E N D
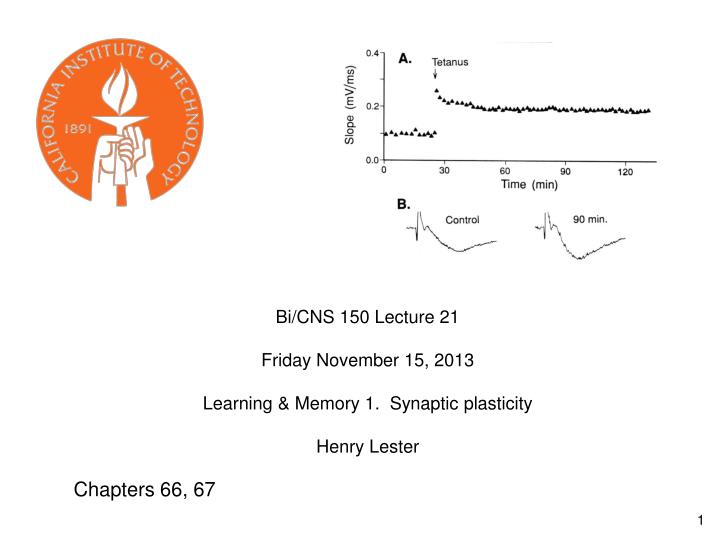
Bi/CNS 150 Lecture 21 Friday November 15, 2013 Learning & Memory 1. Synaptic plasticity Henry Lester Chapters 66, 67
The Hebbian Synapse From The Organization of Behavior by Donald Hebb, 1949: “When an axon of cell A is near enough to excite cell B and repeatedly or persistently takes part in firing it, some growth process or metabolic change takes place in one or both cells such that A's efficiency, as one of the cells firing B, is increased.” Hebb postulated that this behavior of synapses in neuronal networks would permit the networks to store memories. A Hebbian synapse is a “coincidence detector” NMDA receptors, back-propagating action potentials, and summation of epsp’s appear to be the components that confer “Hebbian” behavior on the synapse.
Many synapses occur on spines, which form on dendritic shafts. Spines are dynamic, plastic, changeable. Reconstruction of dendritic spines from serial EM pictures in hippocampus Atlas of Ultrastructural Neurocytology, http://synapse-web.org/atlas/contents.stm
400 nm Electron micrograph of hippocampal synapse “Map” of micrograph to the left
A postsynaptic density, with a cartoon of important proteins AMPA receptor NMDA receptor CaMKII
NMDA receptors are “coincidence detectors”. Their channel opens only when two events happen concurrently: 1. Binding of glutamate • 2. Strong postsynaptic membrane depolarization • (as by an action potential) • The depolarization relieves block by Mg2+ Modified from Zigmond et al. (Eds.) Fundamental Neuroscience, Sinauer (1999) The coincidence readout: NMDA receptors are very permeable to Ca2+
Mechanism of Activation of CaMKII, and “Autophosphorylation” “Ca2+/calmodulin-dependent protein kinase” CaMKII is activated by the calcium-binding protein calmodulin Autophosphorylation of CaMKII can prolong its activation by calcium.
From previous lectures Control of Synaptic Plasticity by NMDA Receptors • The central role of Ca2+ in initiation of long-term plastic changes • The “Ca2+ hypothesis” for control of synaptic plasticity • Measurement of cytosolic Ca2+ with fluorescent dyes. • Control of postsynaptic Ca2+ by “spike timing” • Structure and behavior of the NMDA receptor • Subunit composition • Gating (“coincidence detection”) • Ion selectivity (Na+, K+, Ca2+) • Kinetics, NMDA receptors are slower than AMPA receptors • Pharmacology • The NMDA receptor is also a “scaffold.” • The postsynaptic density • LTP and LTD are triggered by Ca2+-sensitive signaling machinery located near the mouth of the NMDA receptor. • Critical components of the postsynaptic density • Biochemical pathways mediating changes in synaptic strength
Synaptic Plasticity in the Hippocampus, Cortex, and Striatum • Synapses in the cortex and hippocampus are tightly regulated. • Regulation helps to maintain homeostatic balance • 2. It also serves to process and store information in neural circuits. The chemical synapse is a biophysical machine, specialized to function on a time scale of milliseconds and a distance scale < 1 μm. But **also** to adapt to changing needs and activity levels. Homeostasis, adaptation, plasticity, compensation: These are summary processes, not mechanisms. They are best used as adjectives. We do not say, “a synapse changes because of homeostasis”; We say, “a synapse changes because of a homeostatic process” During the 21st century, scientists will continue to discover . . . the molecular instantiations of homeostatic processes.
Presynaptic vs. Postsynaptic • I. The size of synaptic potentials can be modulated: • by regulating the number of number of vesicles (quanta) released. • by regulating the size of the current generated by a released quantum at the postsynaptic membrane. • II. Short term modulation (ms - min) • The mechanisms of these forms of modulation are almost always presynaptic. • Paired-pulse facilitation(~10 to 100 ms) • Synaptic depression(50 ms to min) • Post-tetanic potentiation (min) • Long-term plasticity • The mechanisms of these forms of modulation are usually both pre- and postsynaptic • LTP (30 min to yr) • LTD (30 min to yr)
Paired Pulse Facilitation Paired activations of a synapse onto a Layer 2/3 cortical neuron. “Residual Ca2+” in terminal for 10 to 100 ms after first stimulus increases probability of release.
Short-term Synaptic Depression Successive stimuli at 50 Hz Both the rate and the steady-state level of depression depend on the stimulus frequency. Cook et al. Nature 421, 66-70 (2003)
Post-Tetanic Potentiation PTP believed to be caused by a large accumulation of Ca2+ in the terminal caused by a high frequency tetanic stimulation.
Recording of Long-Term Potentiation in a Hippocampal Slice Stimulation frequencies that produce LTP usually range from ~50 to 200 Hz.
Long-term Synaptic Plasticity • Frequency-dependent Long-term Potentiation (LTP) • This term actually represents many mechanisms, all of which result in strengthening of the synapse for varying periods of time following tetanic stimulation. • The mechanisms for LTP lasting 30 min to a few hr do not require new protein synthesis • The mechanisms for LTP lasting longer than a few hr do require protein synthesis. • Frequency-dependent Long-term Depression (LTD) • This term also represents many mechanisms • LTD, like LTP is thought to be used for sculpting circuits to store information. • Spike-timing dependent synaptic plasticity(STDP) is thought to arise from the same set of mechanisms as LTP and LTD.
Postsynaptic Calcium Levels and Synaptic Plasticity 1. Level and timing of Ca2+ rise in spine determines LTD or LTP. • Low frequency synaptic firing (~5 Hz) produces LTD; • high frequency synaptic firing (~50 to 100 Hz) produces LTP. • The same Ca2+ rules may underlie • “spike-timing-dependent synaptic plasticity (STDP).
Recording of LTD in the Hippocampus Stimulation frequencies usually range from 1 to 10 Hz.
Two cellular processes underlie the major changes during LTP and LTD Insertion of AMPA receptors into the postsynaptic membrane (LTP); or their removal from the postsynaptic membrane (LTD). 2. Growth or shrinkage of the spine via reshaping of the actin cytoskeleton.
LTP is probably “input specific” Figure 67-6
Spike-timing Dependent Synaptic Plasticity Pre- fires 5-30 msbefore post → LTP Pre- fires 5-30 ms after post→LTD These recordings were made on cultured neurons See next slide for experiments in slices. “anti-Hebbian” “Hebbian” From Bi and Poo J. Neurosci. 18, 10464 (1998)
Supralinear influx of Ca2+ during paired EPSP and AP From Schiller, Schiller and Clapham, Nature Neuroscience 1, 114 (1998)
Early LTP, < 2 h Increased size of quantum usually arises from additional receptors Figure 67-8
Late LTP, > 2 h Increased number of quanta usually arises from additional release Protein synthesis is involved Figure 67-8
Overall View of NMDA-dependent LTP in hippocampus (CA3-CA1 synapse) Gene activation via CREB / CRE Dendritic protein synthesis Retrograde signal (nitric oxide?) Cytoskeleton changes?
Role of CaMKII in LTP 1. Calcium ion flows through the activated NMDA receptor. 2. One of its targets is calcium/calmodulin-regulated Protein Kinase II (CaMKII). • 3. CaMKII can phosphorylate the subunits of the AMPA receptor. • The phosphorylated AMPA receptor has a larger current • This is likely one mechanism of relatively short LTP (30 min or so). • 4. CaMKII initiates a process that results in addition of new AMPA receptors to the synapse • This process may be developmentally important • It likely also contributes to longer lasting LTP. 5. Helps regulate processes that re-arrange and enlarge the cytoskeleton.
Role of Calcineurin in LTD 1. Calcium ion flows through the activated NMDA receptor. 2. One of its targets is calcineurin (or protein phosphatase 2B), a Ca2+/CaM-dependent protein phosphatase. • 3. Calcineurin regulates an inhibitor (Inhibitor 1) of a more general protein phosphatase called phosphatase 1. • Inhibition of calcineurin blocks induction of LTD • LTD results from removal of AMPA receptors by endocytosis. • One popular hypothesis is that the direction of long-term changes in synaptic strength depends on the relative levels of activation of CaMKII and calcineurin.
Signals in long-term depression Dominant negative
Distinct Molecular Bases of Long-Term Potentiation at Three Synapses in Hippocampus Figure 67-3
Epigenetic Changes in Chromatin Structure may also Participate in Long-Term Memory Phosphorylation of CREB-1 Recruits CREB Binding Protein (CBP-1) CBP acetylates lysine(+) resides on histones Histones release DNA(-) Allow transcription during late LTP LTP changes DNA methyltransferases (DNMT) This recruits methyl-CpG binding proteins (Me-CpG-BP) This recruits histone deacetylases (HDAC), which remove actyl groups Figure 67-18 This allows CREB-2 binding, which represses transciption Figure 67-18
Henry Lester’s”office” hours 1:15 – 2 M, F Red Door End of lecture 21
Spike-timing Dependent Plasticity in Cortical Neurons Dual whole-cell patch recordings from neurons in cortical slices from 14-16 day old rats (Markram et al., Science 275, 213 (1997)
Record Stim. NMDA-Dependent Long-Term Potentiation in the Hippocampus The third synapse in the “tri-synaptic pathway”