Download

1 / 35
350 likes | 387 Views
Hemopoiesis, the formation of blood cellular components, is crucial for maintaining steady state levels in circulation. This process involves the differentiation of stem cells into various blood cell lineages. Understanding the self-renewal capabilities of hematopoietic stem cells is essential for therapies like bone marrow transplantation. Learn about the different colony-forming units and the importance of the stem cell niche in bone marrow. Discover the role of HSCs in hematopoiesis and their presence in umbilical cord and peripheral blood.

E N D
Haematopoiesis (from Ancient Greek: αἷμα, "blood"; ποιεῖν "to make") (or hematopoiesis in American English; sometimes also haemopoiesis or hemopoiesis) is the formation of blood cellular components. All cellular blood components are derived from haematopoietic stem cells. In a healthy adult person, approximately 1011–1012 new blood cells are produced daily in order to maintain steady state levels in the peripheral circulation.[1][2] http://en.wikipedia.org/wiki/Haematopoiesis
Nagasawa Nature Reviews Immunology6, 107–116 (February 2006) | doi:10.1038/nri1780
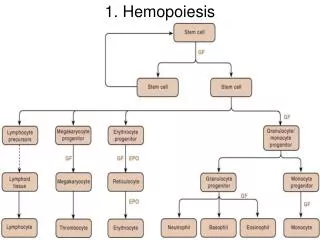
All blood cells are divided into three lineages • Erythroid cells are the oxygen carrying red blood cells. Both reticulocytes and erythrocytes are functional and are released into the blood. In fact, a reticulocyte count estimates the rate of erythropoiesis. • Lymphocytes are the cornerstone of the adaptive immune system. They are derived from common lymphoid progenitors. The lymphoid lineage is primarily composed of T-cells and B-cells (types of white blood cells). This islymphopoiesis. • Myelocytes, which include granulocytes, megakaryocytes and macrophages and are derived from common myeloid progenitors, are involved in such diverse roles as innate immunity, adaptive immunity, and blood clotting. This ismyelopoiesis. • Granulopoiesis (or granulocytopoiesis) is haematopoiesis of granulocytes. • Megakaryocytopoiesis is haematopoiesis of megakaryocytes. http://en.wikipedia.org/wiki/Haematopoiesis
Multipotency and self-renewal • As stem cells, HSC are defined by their ability to replenish all blood cell types (Multipotency) and their ability to self-renew. • It is known that a small number of HSCs can expand to generate a very large number of daughter HSCs. • This phenomenon is used in bone marrow transplantation, when a small number of HSCs reconstitute the hematopoietic system. This process indicates that, subsequent to bone marrow transplantation, symmetrical cell divisions into two daughter HSCs must occur. • Stem cell self-renewal is thought to occur in the stem cell niche in the bone marrow, and it is reasonable to assume that key signals present in this niche will be important in self-renewal. • There is much interest in the environmental and molecular requirements for HSC self-renewal, as understanding the ability of HSC to replenish themselves will eventually allow the generation of expanded populations of HSC in vitro that can be used therapeutically.
There are various kinds of colony-forming units: • Colony-forming unit lymphocyte (CFU-L) • Colony-forming unit erythrocyte (CFU-E) • Colony-forming unit granulo-monocyte (CFU-GM) • Colony-forming unit megakaryocyte (CFU-Me) • Colony-forming unit Basophil (CFU-B) • Colony-forming unit Eosinophil (CFU-Eo) • The above CFUs are based on the lineage. Another CFU, the colony-forming unit–spleen (CFU–S) was the basis of an in vivoclonal colony formation, which depends on the ability of infused bone marrow cells to give rise to clones of maturing hematopoietic cells in the spleens of irradiated mice after 8 to 12 days. It was used extensively in early studies, but is now considered to measure more mature progenitor or Transit Amplifying Cells rather than stem cells.
Haematopoietic stem cells (HSCs) reside in the medulla of the bone (bone marrow) and have the unique ability to give rise to all of the different mature blood cell types and tissues. • HSCs are self-renewing cells: when they proliferate, at least some of their daughter cells remain as HSCs, so the pool of stem cells does not become depleted. • The other daughters of HSCs (myeloid and lymphoid progenitor cells), however can commit to any of the alternative differentiation pathways that lead to the production of one or more specific types of blood cells, but cannot self-renew. This is one of the vital processes in the body. http://en.wikipedia.org/wiki/Haematopoiesis and Hematopoietic_stem_cell
HSCs are also found in umbilical cord blood and, in small numbers, in peripheral blood.Stem and progenitor cells can be taken from the pelvis, at the iliac crest, using a needle and syringe.The cells can be removed a liquid (to perform a smear to look at the cell morphology) or they can be removed via a core biopsy (to maintain the architecture or relationship of the cells to each other and to the bone). • In order to harvest stem cells from the circulating peripheral, blood donors are injected with a cytokine, such as granulocyte-colony stimulating factor (G-CSF), that induce cells to leave the bone marrow and circulate in the blood vessels. • In mammalian embryology, the first definitive HSCs are detected in the AGM (Aorta-gonad-mesonephros), and then massively expanded in the Fetal Liver prior to colonising the bone marrow before birth.[2] http://en.wikipedia.org/wiki/Haematopoiesis and Hematopoietic_stem_cell
In developing embryos, blood formation occurs in aggregates of blood cells in the yolk sac, called blood islands. • As development progresses, blood formation occurs in the spleen, liver and lymph nodes. • When bone marrow develops, it eventually assumes the task of forming most of the blood cells for the entire organism. • Maturation, activation, and some proliferation of lymphoid cells occurs in secondary lymphoid organs (spleen, thymus, and lymph nodes). • In children, haematopoiesis occurs in the marrow of the long bones such as the femur and tibia. In adults, it occurs mainly in the pelvis, cranium, vertebrae, and sternum. • In some cases, the liver, thymus, and spleen may resume their haematopoietic function. This is calledextramedullaryhaematopoiesis. During fetal development, since bones and thus the bone marrow develop later, the liver functions as the main haematopoetic organ. Therefore, the liver is enlarged during development. blood island http://en.wikipedia.org/wiki/Haematopoiesis
Cell Morphology • Exercise • http://www.dartmouth.edu/~anatomy/Histo/lab_4/bonemarrow/DMS104/popup.html http://www.anatomyatlases.org/MicroscopicAnatomy/Section04/Plate0458.shtml
Cell Differentiation • The determinism theory of haematopoiesis, saying that colony stimulating factors and other factors of the haematopoietic microenvironment determine the cells to follow a certain path of cell differentiation. • This is the classical way of describing haematopoiesis. • The ability of the bone marrow to regulate the quantity of different cell types to be produced is more accurately explained by astochastic theory. • Undifferentiated blood cells are determined to specific cell types by randomness. • The haematopoietic microenvironment prevails upon some of the cells to survive and some, on the other hand, to perform apoptosis and die. http://en.wikipedia.org/wiki/Haematopoiesis
Transcription factors • Growth factors initiate signal transduction pathways, altering transcription factors, that, in turn activate genes that determine the differentiation of blood cells. • The early committed progenitors express low levels of transcription factors that may commit them to discrete cell lineages. • Which cell lineage is selected for differentiation may depend both on chance and on the external signals received by progenitor cells. • Several transcription factors have been isolated that regulate differentiation along the major cell lineages. • PU.1 commits cells to the myeloid lineage • GATA-1 has an essential role in erythropoietic and megakaryocytic differentiation. • The Ikaros, Aiolos and Helios transcription factors play a major role in lymphoid development.[5] http://en.wikipedia.org/wiki/Haematopoiesis
The proliferation and self-renewal of these cells depend on stem cell factor (SCF). Glycoproteingrowth factors regulate the proliferation and maturation of the cells that enter the blood from the marrow, and cause cells in one or more committed cell lines to proliferate and mature. • Three more factors that stimulate the production of committed stem cells are called colony-stimulating factors (CSFs) and include granulocyte-macrophage CSF (GM-CSF), granulocyte CSF (G-CSF) andmacrophage CSF (M-CSF). These stimulate much granulocyte formation and are active on either progenitor cells or end product cells. • Erythropoietin is required for a myeloid progenitor cell to become an erythrocyte.[3] • Thrombopoietin makes myeloid progenitor cells differentiate to megakaryocytes (thrombocyte-forming cells).[3] http://en.wikipedia.org/wiki/Haematopoiesis
Nagasawa Nature Reviews Immunology6, 107–116 (February 2006) | doi:10.1038/nri1780
SCF= Stem Cell Factor, Tpo= Thrombopoietin, IL= Interleukin, GM-CSF= Granulocyte Marophage-colony stimulating factor, Epo= Erythropoietin, M-CSF= Macrophage-colony stimulating factor, G-CSF= Granulocyte-colony stimulating factor, SDF-1= Stromal cell-derived factor-1, FLT-3 ligand= FMS-like tyrosine kinase 3 ligand, TNF-a = Tumour necrosis factor-alphaTGF-β = Transforming growth factor beta
Nagasawa Nature Reviews Immunology6, 107–116 (February 2006) | doi:10.1038/nri1780
In this model, the intermediate precursor cells between haematopoeitic stem cells (HSCS) — which are located near the osteoblasts7, 8, endothelial cells113 or CXC-chemokine ligand 12hi (CXCL12hi) reticular cells10 — and pre-pro-B cells would move towards CXCL12hi reticular cells. • Pre-pro-B cells associate with CXCL12hi reticular cells, whereas pro-B cells move away and instead adjoin interleukin-7 (IL-7)-expressing cells10. • Subsequently, pre-B cells leave IL-7-expressing cells10. • B cells expressing cell-surface IgM exit the bone marrow and enter the blood to reach the spleen, where they mature into peripheral mature B cells. • End-stage B cells (plasma cells) again home to CXCL12hi reticular cells in the bone marrow10.
Stem cell heterogeneity • It was originally believed that all HSC were alike in their self-renewal and differentiation abilities. • Muller-Sieburg group in San Diegoillustrated that different stem cells can show distinct repopulation patterns that are epigenetically predetermined intrinsic properties of clonal Thy-1lo SCA-1+ lin- c-kit+ HSC.[3][4][5] • The results of these clonal studies led to the notion of lineage bias. Using the ratio of lymphoid (L) to myeloid (M) cells in blood as a quantitative marker, the stem cell compartment can be split into three categories of HSC. a)Balanced (Bala) HSC repopulate peripheral white blood cells in the same ratio of myeloid to lymphoid cells as seen in unmanipulated mice (on average about 15% myeloid and 85% lymphoid cells, or 3≤ρ≤10). b)Myeloid-biased (My-bi) HSC give rise to too few lymphocytes resulting in ratios 0<ρ<3, c) Lymphoid-biased (Ly-bi) HSC generate too few myeloid cells, which results in lymphoid-to-myeloid ratios of 10<ρ<oo. • All three types are norm three types of HSC, and they do not represent stages of differentiation. Rather, these are three classes of HSC, each with an epigenetically fixed differentiation program
Cluster of differentiation and other markers • Many of markers belong to the cluster of differentiation series, like: CD34, CD38, CD90, CD133, CD105, CD45, and also c-kit, - the receptor for stem cell factor. The hematopoietic stem cells are negative for the markers that are used for detection of lineage commitment, and are, thus, called Lin-; and, during their purification by FACS, a bunch of up to 14 different mature blood-lineage marker, e.g., CD13 & CD33 for myeloid, CD71 for erythroid, CD19 for B cells, CD61 for megakaryocytic, etc. for humans; and, B220 (murineCD45) for B cells, Mac-1 (CD11b/CD18) formonocytes, Gr-1 for Granulocytes, Ter119 for erythroid cells, Il7Ra, CD3, CD4, CD5, CD8 for T cells, etc. (for mice) antibodies are used as a mixture to deplete the lin+ cells or late multipotent progenitors (MPP)s. • There are many differences between the human and mice hematopoietic cell markers for the commonly accepted type of hematopoietic stem cells.[1]. • Mouse HSC :CD34lo/-, SCA-1+, Thy1.1+/lo, CD38+, C-kit+, lin- • Human HSC :CD34+, CD59+, Thy1/CD90+, CD38lo/-, C-kit/CD117+, lin- http://en.wikipedia.org/wiki/Haematopoiesis and Hematopoietic_stem_cell
http://cytometry.nencki.gov.pl/?a=S2vlp8PU http://commons.wikimedia.org/wiki/File:Fluorescence_Assisted_Cell_Sorting_%28FACS%29_A.jpg
Various theories exist about how HSCs diversify • One model (the ‘classical’ model) proposes that lymphocytes and myelo-erythroid lineages branch separately at an early stage of hematopoiesis, • Another model (the ‘myeloid-based’ model) proposes that the myeloid potential is retained for much longer among cells that can become lymphocytes. A revised scheme for developmental pathways of hematopoietic cells: the myeloid-based model International ImmunologyVolume 22, Issue 2Pp. 65-70.
The blood cell family consists of a variety of cell types, all of which are formed from a hematopoietic stem cell (HSC). • Over the last century, the classification of blood cell types was largely based on morphological criteria, leading to the emergence of the classical dichotomy concept, in which the blood cell family was subdivided into two major lineages—a myelo-erythroid lineage and a lymphoid lineage. • Therefore, it has been stated in most textbooks that the first branch point from the HSC produces progenitors for these two lineages.
Representative models of hematopoiesis. (A) HSC firstly generates a common myeloid–erythroid progenitor (CMEP) and a common lymphoid progenitor (CLP), which produce myeloid or erythroid cells and T or B cells, respectively. An alternative myeloid-based model postulates that the HSC first diverges into the CMEP and a common myelo–lymphoid progenitor (CMLP); • (B) In this model, the first branch point generates CMEPs and CMLPs, and the myeloid potential persists in the T and B cell branches even after these lineages have diverged.
The concept of the myeloid-based model. (A) In the classical model, erythroid, myeloid, T and B lineage cells are placed in parallel. (B) The myeloid-based model proposes that myeloid cells represent a prototype of blood cells, whereas erythroid, T and B cells represent specialized types. • Prototypic cells, namely myeloid cells, are equipped with the basic machinery required for host defense cells, e.g. phagocytic activity and mobility. • In the case of B cells, phagocytic activity is reduced but still maintained while the antigen-presenting ability is rather strengthened, and finally, an ability to recognize specific antigen is newly acquired.
T-cell progenitors retain myeloid potential after terminating B cell potential. Early T-cell progenitors in the adult thymus that have lost B-cell potential still retain a substantial capacity to generate macrophages • certain proportion (∼30%) of thymic macrophages are produced by myeloid–T progenitors, by firstly making bone marrow chimeric mice carrying bone marrow cells from wild-type mice and from human-CD3ϵ transgenic mice that lack T lineage cells and subsequently assessing contribution rate of wild-type versus transgenic cells for the production of thymic macrophages (22). • These findings strongly argues against the existence of CLPs on the physiological pathway from the HSC to T cells in adult hematopoiesis.
Schematic illustration of the early differentiation and proliferation of thymic T lineage cells. A single early thymic progenitor undergoes >10 cell divisions during the DN1 and DN2 stages to generate >1000 DN3 cells. The shutoff of myeloid potential occurs during the transition step from the GFP−DN2 stage to the GFP+DN2 stage and subsequently the T-cell lineage-determined progenitors undergo several cell divisions before they enter the DN3 stage to initiate TCRβ chain gene rearrangement.
An illustration of why cell-fate maps should not be over-simplified (using hypothetical cell lineages X, Y, and Z). • (A) An example of the developmental process to produce X cells, Y cells or Z cells. Suppose that a progenitor having potential for X, Y and Z lineages (XYZ-progenitor) first migrates to a particular site (site P); there, it will make X-progenitors and self-renewing XYZ-progenitors, followed by production of X cells from the X-progenitors. • Then, the XYZ-progenitor migrates to the next site (site Q), where they lose their potential to become Y cells to become XZ-progenitors on one hand and on the other hand segregation to Y-progenitors also occurs that become Y cells. • Note that the XZ progenitors do not produce X cells in site Q but can produce X cells in other place. The XZ-progenitor then migrates to site R and produces Z-progenitors and finally Z cells there.
A simplified model for the process shown in (A), which contains information about developmental potential and cell fate. A map like this is useful not only to understand reality but also for further investigations into differentiation mechanisms. • A map of lineage restriction focusing on the way from the XYZ-progenitor to a Z cell. Particularly in studying the molecular mechanisms in lineage commitment for the production of Z cells, the information for the order of lineage restriction [XYZ → XZ → Z] is essential.
A map that describes only the physiological cell fate. This map might be misleading because the information about the lineage restriction process shown in (C) is absent.