Download

1 / 33
1.57k likes | 7.92k Views
Gastrulation. Definition = migration and division of cells to set up the 3 primary germ layers . What positions do presumptive germ layers occupy in the blastula, before gastrulation ?

E N D
Gastrulation • Definition = migration and division of cells to set up the 3 primary germ layers. • What positions do presumptive germ layers occupy in the blastula, before gastrulation? • Experimental answers to this question provide our basis for understanding cell movements that occur during gastrulation. • FATE MAPS = diagram of blastula/blastodisc showing the “fate” of each part. • Fate mapping technique developed by Vogt in 1920s. Involves … • Marking surface of blastula with vital dyes • Dyes retained by cells for prolonged periods, but don’t interfere with normal cellular processes • Follow movements of marked cells during gastrulation to ultimate locations in later embryos
Gastrulation • Micro- and Mesolecithal eggs (Amphioxus and Amphibians) … • Basic cell movements during gastrulation are spreading and stretching, accompanied by cell division. • 7 stages to gastrulation in micro- and mesolecithal eggs.
Steps in Gastrulation • Prospective endodermal cells in vegetal hemisphere elongate and move toward interior. • Base of cell remains attached to surface of blastula, this causes strain on surface which causes an indentation to form (blastopore). • Area directly above the blastopore = Dorsal Lip of Blastopore. This is where cells begin to move inward. • Invagination spreads circulolaterally enclosing yolk plug of endoderm. • Continued invagination and spreading so that surface of gastrula contains only ectoderm. • Notochord and somatic mesoderm become stretched out longitudinally inside, neural ectoderm longitudinally on surface. • When complete: • composite inner layer involving both mesoderm and endoderm • notochordal mesoderm and neural ectoderm in parallel • blastocoel eliminated by spreading of endoderm • yolk plug has withdrawn internally so blastopore is only a narrow slit
Fig 5.7 – Gastrulation and Neurulation in Amphioxus
Fig 5.11 – Gastrulation and Neurulation in Amphibians
Amphibian Gastrulation Video http://worms.zoology.wisc.edu/frogs/gastxen/wholegas.html
Gastrulation in Macrolecithal Eggs • Pattern of movements is different, but the process is basically the same. • Elasmobranchs = basically infolding over posterior margin of blastodisc. • Outer surface becomes ectoderm, inner surface endoderm, mesoderm resides in between. • Entire embryo still on top of yolk.
Gastrulation in Macrolecithal Eggs Reptiles and Birds: • Blastodisc contains cells of varying size: large, yolk-rich cells accumulate at lower and posterior surface, small yolk-poor cells at upper surface. • A separation gradually forms between large lower and small upper cells (= blastocoel), forms epiblast(dorsally) and hypoblast (ventrally). • Upper epiblast forms prospective ectoderm and mesoderm, lower hypoblast forms prospective endoderm. • Hypoblast spreads anterolaterally to form endoderm layer. • Mesoderm infolding occurs in central region of embryo. • Infolding begins at posterior end of blastodisc; progresses anteriorly forming a groove bounded by parallel ridges = primitive streak • At anterior end of this groove, there exists a raised node of tissue with a pit (primitive pit) extending down and forward beneath it. Node = Henson’s Node – equivalent of dorsal lip of blastopore in micro- and mesolecithal eggs. • Mesodermal tissue migrates downward along primitive streak expanding laterally beneath ectoderm and above endoderm.
Fig 5.13 – Gastrulation in the Bird embryo
Chick Gastrulation Video http://www.gastrulation.org/
Gastrulation in Macrolecithal Eggs • Hypoblast may exert causal influence on primitive streak: • If hypoblast separated from epiblast and reoriented, the axis of the primitive streak is similarly reoriented. • If hypoblast removed, primitive streak fails to form • These experiments suggest that the hypoblast forms a necessary substratum for orientation and deployment of prospective mesoderm.
Gastrulation in Mammals • Cavities form above and below ICM, which expand (as amnion and yolk sac, respectively), leaving a flat 2-layered plate of cells. • Primitive streak forms as in birds and reptiles to produce primary germ layers.
Major Tissue Regions after Gastrulation • Skin ectoderm • Neural ectoderm • Notochordal mesoderm • Lateral mesoderm • Endoderm
NEURAL TUBE FORMATION – Amphioxus • Folding up of tissue at junction of future skin ectoderm and neural ectoderm areas; the two tissues separate as this fold forms • Skin ectoderm grows over the top of neural ectoderm • Beneath “skin,” lateral margins of neural ectoderm grow upward and together to form tube • Tube first closes at midpoint, progresses anteriorly and posteriorly. Anterior end opens to surface as neuropore, posterior end forms common opening with blastopore (becomes anus) forming neurenteric canal.
Fig 5.7 – Gastrulation and Neurulation in Amphioxus
NEURAL TUBE FORMATION – Vertebrates • Formation of neural folds along margins of skin-neural ectoderm • Mid-dorsal meeting of folds, simultaneous with joining of skin ectoderm • During folding, high crests of tissue are formed on either side = neural crest cells
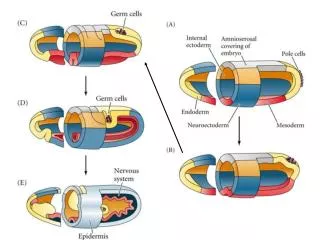
Fig 5.16 – Gastrulation and Neurulation in Mammals
MESODERM DEVELOPMENT • Majority of body structures are mesodermal in origin. • NotochordalMesodermrapidly rounds up and separates from lateral mesoderm, forming a discrete cylinder = notochord. • Notochord is much reduced or obliterated in most adult vertebrates, but forms the center around which vertebral formation occurs. • Lateral Mesoderm– Amphioxus • Mesoderm forms paired series of segmentally arranged blocks = somites. • From their initiation, somites have a cavity inside = coelomic cavity.
MESODERM DEVELOPMENT • Lateral Mesoderm– Vertebrates • Initially there is no segmentation of mesoderm; instead forms as a continuous sheet without a central cavity. • Mesodermal differentiation occurs from dorsal midline outward into 3 divisions, each extending the entire length of the body trunk. • Differentiation always occurs head-to-tail. • The 3 divisions are: • Next to neural tube and notochord = Epimere (somites). Thicken and subdivide on either side to form longitudinal rows of blocks. This is the first indication of segmentation in vertebrate embryos. Proliferation and differentiation occurs within somite forming: • Sclerotome = portion surrounding notochord and neural tube • Dermatome = outermost portion near skin ectoderm • Myotome = middle portion between and ventral to sclerotome and dermatome
MESODERM DEVELOPMENT • Lateral and ventral to somites is a relatively small region of mesoderm, known as intermediate mesoderm or Mesomere. This may show segmentation similar to somites. • Beyond mesomere region, extending ventrolaterally is a sheet of mesoderm known as lateral plate mesoderm or Hypomere. • Apart from cyclostomes, there is no segmentation in this region. Coelomic cavity forms within lateral plate mesoderm, dividing it into: • Somatopleure= external mesoderm + ectoderm • Splanchnopleure = internal mesoderm + endoderm
Figs 5.11 & 5.16 Mesoderm divisions in Amphibians and Mammals
Organogenesis/Differentiation • Once the mesoderm divisions are set up, then ontogenetic development proceeds to embryonic differentiation to adult body. • What causes this differentiation? • Induction = process by which developmental fate of cells is determined
Induction • Classic Experiments of Spemann (1920s) – won Nobel Prize • Took piece of presumptive neural plate from early gastrula stage, transplanted to ventral region of another early gastrula normal development, prospective neural ectoderm becomes skin ectoderm. • Similar experiment with late gastrula transplanted neural ectoderm becomes neural ectoderm regardless of transplant site. • Transplant dorsal lip (prospective notochordal mesoderm) of early gastrula to ventral region in another embryo induces formation of a secondary embryo involving both host and transplanted tissues. • Conclusions: • Some change took place between early and late gastrula that determined fate of cells • Dorsal lip responsible for inducing shift in direction of differentiation of host tissue. Dorsal lip plays a central role in determining the craniocaudal axis of the embryo = Primary Organizer.
Spemann Expt - Transplanted dorsal lip (prospective notochordal mesoderm) of early gastrula to ventral region in another embryo induces formation of a secondary embryo involving both host and transplanted tissues.
Primary Induction • Definition = induction with primary importance in determining cranio-caudal axis, also the first of many inductive interactions. • Two types of inductive interactions: • Instructive = directs differentiation of cells along a certain path • Permissive = allows differentiation when given a proper stimulus
What is the Mechanism of Induction? • Classic Triturus(Salamander) experiment … • Dorsal Lip (Inducer) cultured for 24 hr across nitrocellulose membrane with pores of specific diameter from undifferentiated ectoderm →Ectoderm differentiates to become neural tissues. • If undifferentiated ectoderm cultured in the absence of dorsal lip, it develops into unspecialized skin ectoderm. • EM analysis of membrane after culture showed no cellular processes between dorsal lip and ectoderm. • Conclusion = a diffusable substance responsible for induction.
What is the Mechanism of Induction? • Chemical nature of diffusable substance? • Not known with certainty • Purified active ingredients from various inducers turn out to be proteins or glycoproteins • Also some evidence that changes in ratio of bound/free ions within cells of early gastrula may influence induction.
What is the Mechanism of Induction? • Other inductive interactions between cells can result from … • Cell-to-cell direct physical interactions – usually between molecules located on the cell surface • Cell-to-cell communication of signals through gap junctions • For induction via diffusable substances or direct physical interactions, cell membrane receptors on the induced cells are required
SUMMARY • The full developmental pathway is dependent upon the genetic and biochemical capacities of the induced cells + the full inductive capabilities of the inducing tissue. • Many inductive interactions occur throughout development and influence gene expression and the production of specific gene products and cell migration, among other processes • Precise mechanisms remain uncertain in most cases.