Download
1 / 34
420 likes | 777 Views
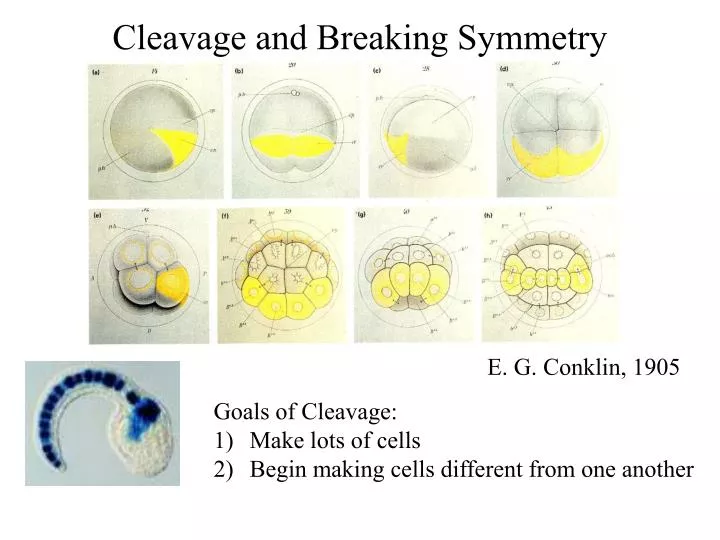
Cleavage and Breaking Symmetry. E. G. Conklin, 1905. Goals of Cleavage: Make lots of cells Begin making cells different from one another. Intrinsic Mechanisms. How to give daughter cells DIFFERENT Identities. Extrinsic Mechanisms. Autonomous specification

E N D
Cleavage and Breaking Symmetry E. G. Conklin, 1905 Goals of Cleavage: Make lots of cells Begin making cells different from one another
Intrinsic Mechanisms How to give daughter cells DIFFERENT Identities Extrinsic Mechanisms
Autonomous specification Cell fate determined by intrinsic mechanism (e.g. cytoplasmic determinant) Mosaic Development Cells of embryo retain discrete fates when separated (e.g. Ascidian embryo) The “European Plan”: You develop according to where you were born and lineage is important. (after S. Brenner) As opposed to- Conditional specification Cell fate determined by extrinsic mechanism (e.g. signalling) Regulative Development Cells of embryo can modify cell fate to form new structures when isolated (e.g. human identical twins) The “American Plan”: Cells start off undetermined, albeit with some biases. They are determined by their environment, sometimes after migrating long distances, and tend to behave similarly to their neighbors. (after S. Brenner)
Cytoplasmic Rearrangement in Ascidian Zygote A) Before fertilization Uniform cortical yellow cytoplasm Gray, yolky cytoplasm inside B) After sperm entry Yellow cytoplasm streams towards sperm nucleus C and D) Yellow cytoplasm moves with migrating sperm nucleus Yellow crescent is formed
Fate Mapping the Ascidian Embryo Yellow Crescent E. G. Conklin, 1905
Blastomere Isolation Experiments Muscle No Muscle Whittaker, 1977 -Blastomere pair containing yellow crescent can form muscle when cultured in isolation -Muscle markers come on at appropriate time relative to control embryos -Muscle markers come on even when cell division is blocked -Remainder of embryo does not form muscle
The Ultimate in Blastomere Isolation: Nishida (Roux’s Archives, 1992) -Separate blastomeres after each division until 110 cell stage -Allow blastomeres to develop separately -Get many individual cell fates: muscle, gut, epidermis BUT NOT: notochord or nervous system Thus: muscle is AUTONOMOUSLY SPECIFIED notochord is CONDITIONALLY SPECIFIED
Asymmetric cell division segregates cytoplasmic determinants Need to: 1) Break Symmetry 2) Establish cell polarity 3) Localize cytoplasmic determinants 4) Control mitotic spindle
Asymmetric segregation of P granules to P cell lineage Susan Strome
C. elegans first cleavage asymmetries: 1) Cytoplasmic flow toward posterior P granules/PIE-1 at posterior Asymmetric cleavage (post. cell smaller) Different cell fates P Granules (Stome Lab) PIE-1-GFP (Seydoux Lab) Goldstein Lab
Asymmetric Cell Division 1) Break Symmetry 2) Establish Cell Polarity 3) Localize Determinants 4) Control Mitotic Spindle Gonczy Nature Reviews, 2008
Sperm Entry Point Determines Posterior in C. elegans (Goldstein and Hird)
Sadler and Shakes wt No sperm nucleus The paternal pronucleus is not required for A/P patterning in C. elegans
Laser ablation of centrosomes disrupts polarity Time of ablation (relative to establishment of polarity) after before during GFP-PAR2
Breaking Symmetry: It all starts at the cortex 1) Sperm contributes centrosomes and Rho Gap 2) Creates asymmetry in cortical domains 3) Actin/myosin contractility more active at anterior than posterior 4) Helps move “anterior” PARs to anterior and allow “posterior” PARs to localize Nance, 2005 Goldstein Lab
Asymmetric Cell Division 1) Break Symmetry 2) Establish Cell Polarity 3) Localize Determinants 4) Control Mitotic Spindle Gonczy Nature Reviews, 2008
Cell Polarity: Specifying Sub-Cellular Identity PAR6-GFP PAR2-GFP
A conserved machinerly for cell polarity Polarized epithelium Polarized zygote
Anterior Complex PAR3 PDZ Domain (prot-prot) PAR6 PDZ Domain PKC3 Atypical Prot. Kinase C Posterior Complex PAR2 Ring Finger PAR1 Ser/Thr Kinase Not Localized PAR4 Ser/Thr Kinase PAR5 14-3-3 Who needs who for localization: Anterior complex required for initial localization of PAR2 (PKC3 inhibits phosphorylates and blocks PAR2) PAR2 NOT required for initial localization of anterior complex PAR2 required for maintenance of anterior complex (phos. of PAR3 by PAR1?) PAR1 is most downstream: dependent on all others for localization
Asymmetric Cell Division 1) Break Symmetry 2) Establish Cell Polarity 3) Localize Determinants 4) Control Mitotic Spindle Gonczy Nature Reviews, 2008
Segregation of Cell Fate Determinants Pie-1-GFP Priess Lab P Granules Strome Lab Seydoux Lab
Centrosomes Actin/myosin ZIF-1 Dependent Degradation
Asymmetric Cell Division 1) Break Symmetry 2) Establish Cell Polarity 3) Localize Determinants 4) Control Mitotic Spindle Gonczy Nature Reviews, 2008
Linking Cell Polarity to Spindle Orientation: A Conserved Model Cell Polarity MT Motor /Par-3 Connect Cell Polarity to Motor NuMA-type Dynein Binder “GoLoco/TPR” Protein Non-receptor Heterotrimeric G Protein -Par complex localizes GPR1/2 -G-alpha activates GPR1/2 -GPR1/2 bind dynein complex -dynein complex orients spindle Worm Fly Mouse Siller and Doe, Nature Cell Biol. 2009
Spindle Dynamics in Early C. elegans Zygote GPR-1/2 at Anterior GPR-1/2 at Posterior Siller and Doe, Nature Cell Biol. 2009
The posterior “pulls harder” on the spindle to generate different size daughter cells
Asymmetric Cell Division 1) Break Symmetry 2) Establish Cell Polarity Localize Determinants 4) Control Mitotic Spindle
PAR-1 PAR-1 PAR-3-GFP S-A PAR-3-GFP wt PAR-3-GFP STAU PAR-3-GFP PAR-3 PAR-1 PARs and Oocyte Patterning in Drosophila St. Johnston Lab
Asymmetric Cell Division in Neuroblasts Apical Complex e.g. Baz/Par3 Cell Fate Determinants e.g. Miranda, Prospero Baz = Par3 Bellaiche and Gotta, 2005 Wodarz, 2005