Download
1 / 1
10 likes | 148 Views
Can Microtubule Dynamics Be Explained Using the Geometry Characteristic of the B-Lattice?. Maria J. Schilstra 1 , John J. Correia 2 , and Stephen R.Martin 3 1 STRI, University of Hertfordshire, College Lane, Hatfield AL10 9AB, UK; m.j.1.schilstra@herts.ac.uk;

E N D
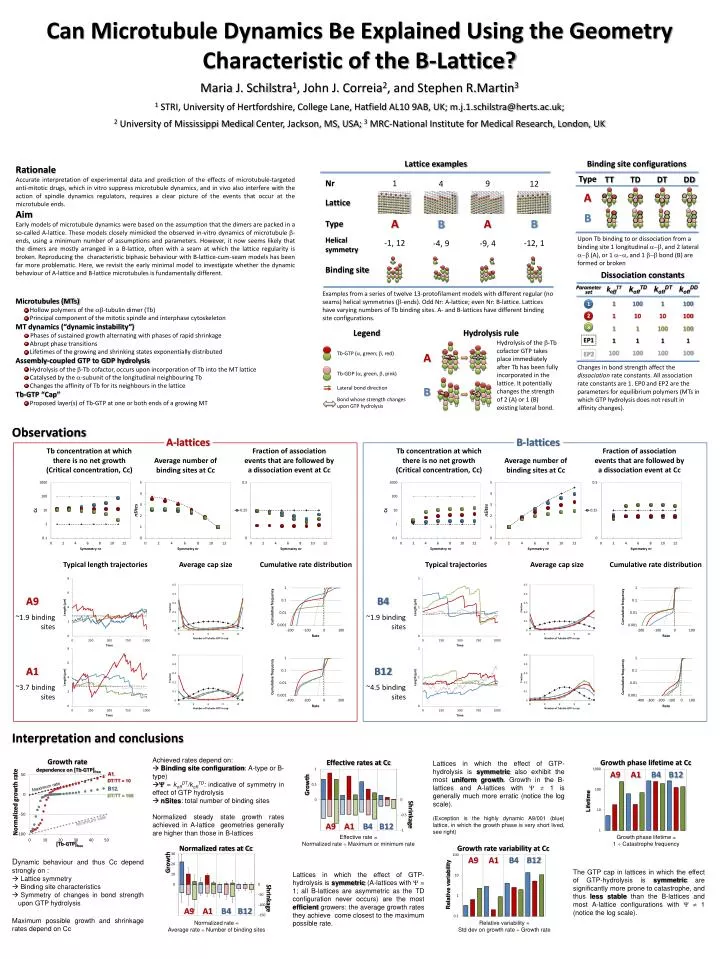
Can Microtubule Dynamics Be Explained Using the Geometry Characteristic of the B-Lattice? Maria J. Schilstra1, John J. Correia2, and Stephen R.Martin3 1 STRI, University of Hertfordshire, College Lane, Hatfield AL10 9AB, UK; m.j.1.schilstra@herts.ac.uk; 2 University of Mississippi Medical Center, Jackson, MS, USA; 3 MRC-National Institute for Medical Research, London, UK Rationale Accurate interpretation of experimental data and prediction of the effects of microtubule-targeted anti-mitotic drugs, which in vitro suppress microtubule dynamics, and in vivo also interfere with the action of spindle dynamics regulators, requires a clear picture of the events that occur at the microtubule ends. Aim Early models of microtubule dynamics were based on the assumption that the dimers are packed in a so-called A-lattice. These models closely mimicked the observed in-vitro dynamics of microtubule -ends, using a minimum number of assumptions and parameters. However, it now seems likely that the dimers are mostly arranged in a B-lattice, often with a seam at which the lattice regularity is broken. Reproducing the characteristic biphasic behaviour with B-lattice-cum-seam models has been far more problematic. Here, we revisit the early minimal model to investigate whether the dynamic behaviour of A-lattice and B-lattice microtubules is fundamentally different. Lattice examples Binding site configurations Type TT TD DT DD Lattice A 1 Nr 9 12 4 B A A B B Type Upon Tb binding to or dissociation from a binding site 1 longitudinal a-b, and 2 lateral a-b (A), or 1 a-a, and 1 b-b bond (B) are formed or broken Helical symmetry -1, 12 -12, 1 -9, 4 -4, 9 Binding site Dissociation constants koffTT koffTD koffDT koffDD Parameter set Examples from a series of twelve 13-protofilament models with different regular (no seams) helical symmetries (-ends). Odd Nr: A-lattice; even Nr: B-lattice. Lattices have varying numbers of Tb binding sites. A- and B-lattices have different binding site configurations. • Microtubules (MTs) • Hollow polymers of the -tubulin dimer (Tb) • Principal component of the mitotic spindle and interphase cytoskeleton • MT dynamics (“dynamic instability“) • Phases of sustained growth alternating with phases of rapid shrinkage • Abrupt phase transitions • Lifetimes of the growing and shrinking states exponentially distributed • Assembly-coupled GTP to GDP hydrolysis • Hydrolysis of the b-Tb cofactor, occurs upon incorporation of Tb into the MT lattice • Catalysed by the -subunit of the longitudinal neighbouring Tb • Changes the affinity of Tb for its neighbours in the lattice • Tb-GTP “Cap” • Proposed layer(s) of Tb-GTP at one or both ends of a growing MT 1 100 1 100 1 1 10 10 100 2 1 1 100 100 3 Legend Hydrolysis rule EP1 1 1 1 1 Hydrolysis of the b-Tb cofactor GTP takes place immediately after Tb has been fully incorporated in the lattice. It potentially changes the strength of 2 (A) or 1 (B) existing lateral bond. 100 100 100 100 Tb-GTP (a, green; b, red) EP2 A Changes in bond strength affect the dissociation rate constants. All association rate constants are 1. EP0 and EP2 are the parameters for equilibrium polymers (MTs in which GTP hydrolysis does not result in affinity changes). Tb-GDP (a, green, b, pink) B Lateral bond direction Bond whose strength changes upon GTP hydrolysis Observations B-lattices A-lattices Tb concentration at which there is no net growth (Critical concentration, Cc) Fraction of association events that are followed by a dissociation event at Cc Average number of binding sites at Cc Typical length trajectories Average cap size Cumulative rate distribution A9 ~1.9 binding sites Typical trajectories Average cap size Cumulative rate distribution A1 ~3.7 binding sites B4 ~1.9 binding sites Tb concentration at which there is no net growth (Critical concentration, Cc) Fraction of association events that are followed by a dissociation event at Cc Average number of binding sites at Cc B12 ~4.5 binding sites Interpretation and conclusions • Achieved rates depend on: • Binding site configuration: A-type or B-type) • = koffDT/koffTD: indicative of symmetry in effect of GTP hydrolysis • nSites: total number of binding sites • Normalized steady state growth rates achieved in A-lattice geometries generally are higher than those in B-lattices Growth rate dependence on [Tb-GTP]free Lattices in which the effect of GTP-hydrolysis is symmetric also exhibit the most uniform growth. Growth in the B-lattices and A-lattices with 1 is generally much more erratic (notice the log scale). (Exception is the highly dynamic A9/001 (blue) lattice, in which the growth phase is very short lived, see right) A1, DT/TT = 10 Maximum rate B12, DT/TT = 100 Normalized growth rate Minimum rate [Tb-GTP]free Normalized rates at Cc Effective rates at Cc Growth rate variability at Cc Growth phase lifetime at Cc • Dynamic behaviour and thus Cc depend strongly on : • Lattice symmetry • Binding site characteristics • Symmetry of changes in bond strength upon GTP hydrolysis • Maximum possible growth and shrinkage rates depend on Cc A9 A1 B4 B12 A9 A1 B4 B12 Growth The GTP cap in lattices in which the effect of GTP-hydrolysis is symmetric are significantly more prone to catastrophe, and thus less stable than the B-lattices and most A-lattice configurations with 1 (notice the log scale). Lattices in which the effect of GTP-hydrolysis is symmetric (A-lattices with = 1; all B-lattices are asymmetric as the TD configuration never occurs) are the most efficient growers: the average growth rates they achieve come closest to the maximum possible rate. Growth Relative variability Lifetime Shrinkage Shrinkage A9 A1 B4 B12 A9 A1 B4 B12 Normalized rate = Average rate Number of binding sites Effective rate = Normalized rate Maximum or minimum rate Relative variability = Std dev on growth rate Growth rate Growth phase lifetime = 1 Catastrophe frequency






















