Download
1 / 1
10 likes | 107 Views
The influence of polymorphism for Regulator of G-protein signaling 4 (RGS4) Gene on Regional Brain Metabolism ( 18 FDG PET) and phenotypes in schizophrenia. J . Hor ac ek, F . Spaniel, M. Kopecek, T . Novak, P. Mohr, C. Höschl

E N D
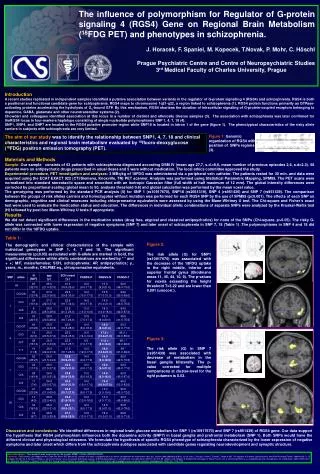
The influence of polymorphism for Regulator of G-protein signaling 4 (RGS4) Gene on Regional Brain Metabolism (18FDG PET) and phenotypes in schizophrenia. J. Horacek, F. Spaniel, M. Kopecek, T.Novak, P. Mohr, C. Höschl Prague Psychiatric Centre and Centre of Neuropsychiatric Studies 3rd Medical Faculty of Charles University, Prague Introduction A recent studies replicated in independent samples identified a putative association between variants in the regulator of G-protein signalling 4 (RGS4) and schizophrenia. RGS4 is both a positional and functional candidate gene for schizophrenia. RGS4 maps to chromosome 1q21-q22, a region linked to schizophrenia (1). RGS4 protein functions primarily as GTPase-activating proteins accelerating the hydrolysis of Gα-bound GTP. By this mechanism, RGS4 shortens the duration of intracellular signaling of G-protein-coupled receptors belonging to dopamine, GABA, glutamate and other neurotransmitter systems (2). Chowdari and colleagues identified association at this locus in a number of distinct and ethnically diverse samples (3). The association with schizophrenia was later confirmed for theRGS4 locus in four-markers haplotype consisting of single nucleotide polymorphisms SNP 1, 4, 7, 18 (4). SNP1, SNP4, and SNP7 are located in the RGS4 putative promoter region while SNP18 is located in intron 1 of the gene (figure 1). The phenotypical characteristics of the risky allele carriers in subjects with schizophrenia are very limited. The aim of our study was to identify the relationship between SNP1, 4, 7, 18 and clinical characteristics and regional brain metabolism evaluated by 18fluoro-deoxyglucose (18FDG) positron emission tomography (PET). Figure 1: Genomic organization of RGS4 with position of SNPs regions (3). Materials and Methods Sample: Our sample consists of 63 patients with schizophrenia diagnosed according DSM IV (mean age 27,7, s.d.=8,6, mean number of previous episodes 2,6, s.d=2.3). 58 patients were on antipsychotic drugs prescribed in usual doses and 5 were without medication. The local ethics committee approved the study. Experimental procedure, PET investigation and analyses: 3 MBq/kg of 18FDG was administered via a peripheral vein catheter. The patients rested for 30 min. and data were acquired using the ECAT EXACT 922 (CTI/Siemens, Knoxville, TN) PET scanner. Analysis was performed using Statistical Parametric Mapping, SPM99. The PET scans were normalized into standard stereotactic space and smoothed with an isotropic Gaussian filter (full width at half maximum of 12 mm). The global intensity differences were corrected by proportional scaling (global mean to 50, analysis threshold 0.8) and global calculation was performed by the mean voxel value. The genotyping was performed by the standard PCR analysis (5) for SNP 1 (rs10917670), SNP18 (rs2661319), SNP 4 (rs951436) and SNP 7 (rs951439). The comparison between the groups of risk alleles carriers and homozygotes without mutation was performed for each RGS4 SNP by the use of SPM99 (p0.001). The effects of each SNP on demographic, cognitive and clinical measures including chlorpromazine equivalents were assessed by using the Mann-Whitney U test. The Chi-square and Fisher’s exact test were used to evaluate the medication status and education. The differences in individual allelic combinations of separate SNPs were analyzed by the Kruskal-Wallis test (H) followed by post hoc Mann-Whitney U tests if appropriate. Results We did not detect significant differences in the medication status (drug free, atypical and classical antipsychotics) for none of the SNPs (Chi-square, p>0.05). The risky G-allele was connected with lower expression of negative symptoms (SNP 7) and later onset of schizophrenia in SNP 7, 18 (Table 1) .The polymorphisms in SNP 4 and 18 did not differ in the 18FDG uptake. Table 1: The demographic and clinical characteristics of the sample with individual genotypes in SNP 1, 4, 7 and 18. The significant measurements (p0.05) associated with G-allele are marked in bold, the significant differences within allelic combinations are marked by * * and + +; M/F, males/females; SCH, schizophrenia; AP, antipsychotics; y., years; m., months; CHLPMZ eq., chlorpromazine equivalents. Figure 2: The risk allele (G) for SNP1 (rs10917670) was associated with the decrease of the 18FDG uptake in the right middle, inferior and superior frontal gyrus (Brodmann areas 11, 45, 44, 10, 9). The p-values for voxels exceeding the height threshold T=3.22 and are lower than 0.001 (unocorr.). Figure 3: The risk allele (G) in SNP 7 (rs951439) was associated with decrease of metabolism in the basal ganglia bilaterally. The p-value corrected for multiple comparisons at cluster-level for the right putamen is 0.03. Discussion and conclusions: We identified differences in regional brain glucose metabolism for SNP 1 (rs10917670) and SNP 7 (rs951439) of RGS4 gene. Our data support the hypothesis that RGS4 polymorphism influences both the dopamine activity (SNP7) in basal ganglia and prefrontal metabolism (SNP 1). Both SNPs would have the different clinical and physiological relevance.We formulate the hypothesis of specific RGS4 phenotype of schizophrenia characterized by the lower expression of negative symptoms and later onset which differs from the schizophrenia subtypes associated with candidate genes regulating neurodevelopment and synaptic structure. Acknowledgment: The research was supported by the projectMSMT CR No.1M0002375201. References:(1)Brzustowicz LM, Hodgkinson KA, Chow EW, Honer WG, Bassett AS. Location of a major susceptibility locus for familial schizophrenia on chromosome 1q21-q22. Science 2000;288(5466):678-82. (2) De Vries L, Zheng B, Fischer T, Elenko E, Farquhar MG. The regulator of G protein signaling family. Annu.Rev.Pharmacol.Toxicol. 2000;40:235-71. (3) Chowdari KV, Mirnics K, Semwal P, Wood J, Lawrence E, Bhatia T et al. Association and linkage analyses of RGS4 polymorphisms in schizophrenia. Hum.Mol.Genet. 2002;11(12):1373-80. (4)Morris DW, Rodgers A, McGhee KA, Schwaiger S, Scully P, Quinn J et al. Confirming RGS4 as a susceptibility gene for schizophrenia. Am J Med.Genet. 2004;125B(1):50-3. (5) Minarik M, Minarikova L, Bjorheim J, Ekstrom PO. Cycling Gradient Capillary Electrophoresis (CGCE): A low cost tool for high-throughput analysis of genetic variations, Electrophoresis, 2003; (24): 1716-1722.






