Download

1 / 30
730 likes | 2.87k Views
Carbohydrate Metabolism 2: Glycogen degradation, glycogen synthesis, reciprocal regulation of glycogen metabolism. Bioc 460 Spring 2008 - Lecture 34 (Miesfeld). Glycogen phosphorylase enzyme is a dimer that is regulated by both phosphorylation and allostery.

E N D
Carbohydrate Metabolism 2:Glycogen degradation, glycogen synthesis, reciprocal regulation of glycogen metabolism Bioc 460 Spring 2008 - Lecture 34 (Miesfeld) Glycogen phosphorylase enzyme is a dimer that is regulated by both phosphorylation and allostery Carbohydrates in pasta are a good way to replenish muscle glycogen stores Gerty Cori won the 1947 Nobel Prize for her work on glycogen metabolism
Key Concepts in Glycogen Metabolism • Glycogen is a highly-branched polymer of glucose that can be quickly degraded to yield glucose-1P which is isomerized to glucose-6P for use in glycolysis by muscle cells, or it is dephosphorylated in liver cells and exported. • Glycogen phosphorylase removes one glucose at a time from the nonreducing ends using inorganic phosphate (Pi) which makes glucose release a “free” reaction. Glycogen phophorylase is activated by phosphorylation in response to glucagon and epinephrine, and allosterically-regulated by energy charge. • Glycogen synthase adds glucose residues to nonreducing ends in a reaction involving UDP-glucose, a nucleotide charged form of glucose. Since the UDP that is released following glucose addition needs to be phosphorylated to regenerate UTP, the cost of glycogen synthesis is 1 ATP/glucose residue. • Glycogen phosphorylase and glycogen sythase are reciprocally-regulated by phosphorylation. Net phosphorylation leads to glycogen degradation, whereas, net dephosphoryation, results in glycogen synthesis.
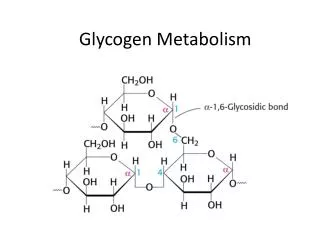
Overview of Glycogen Metabolism The storage form of glucose in most eukaryotic cells (except plants) is glycogen, a large highly branched polysaccharide consisting of glucose units joined by -1,4 and -1,6 glycosidic bonds. Most glycogen in animals is stored in the muscle and liver tissues. Glycogen degradation and synthesis occurs in the cytosol and the substrate for these reactions is the free ends of the branched polymer (nonreducing ends). The large number of branch points in glycogen results in the generation of multiple nonreducing ends that provide a highly efficient mechanism to quickly release and store glucose.
The reducing and nonreducing ends of glycogen The nonreducing end of glycogen molecules refers to the carbon that is opposite to the reducing end in the ring structure. The reducing end of a linear glucose molecule can be oxidized by Cu2+ by definition. Reducing end Nonreducing end Nonreducing end ……… Reducing end
Glycogen Core Complexes Glycogen core complexes consist of glycogenin protein and ~50,000 glucose molecules with α-1,6 branches about every 10 residues creating ~2,000 nonreducing ends. Twenty to forty glycogen core complexes associate inside liver and muscle cells to form glycogen particles containing over a million glucose molecules. These glycogen particles can be visualized by electron microscopy and account for up to 10% by weight of liver tissue.
Pathway Questions Liver glycogen is used as a short term energy source for the organismby providing a means to store and release glucose in response to blood glucose levels; liver cells do not use this glucose for their own energy needs. Muscle glycogen provides a readily available source of glucose during exercise to support anaerobic and aerobic energy conversion pathways within muscle cells; muscle cells lack the enzyme glucose-6-phosphatase and therefore cannot release glucose into the blood.
Pathway Questions 2. What are the net reactions of glycogen degradation and synthesis? Glycogen Degradation: Glycogenn units of glucose + Pi→ Glycogenn-1 units of glucose + glucose-6-phosphate Glycogen Synthesis: Glycogenn units of glucose + glucose-6-phosphate + ATP + H2O → Glycogenn+1 units of glucose + ADP + 2Pi
Pathway Questions 3. What are the key enzymes in glycogen metabolism? Glycogen phosphorylase – enzyme catalyzing the phosphorylysis reaction that uses Pi to remove one glucose at a time from nonreducing ends of glycogen resulting in the formation of glucose-1P. Liver and muscle glycogen phosphorylase are isozymes (two different genes) that are both activated by phosphorylation but have distinct responses to allosteric effectors.Glycogen synthase - enzyme catalyzing the addition of glucose residues to nonreducing ends of glycogen using UDP-glucose as the glucose donor. Glycogen synthase activity is inhibited by phosphorylation; binding of the allosteric activators glucose or glucose-6P promotes dephosphorylation and enzyme activation.Branching and debranching enzymes - these two enzymes are responsible for adding (branching) and removing (debranching) glucose residues to the glycogen complex through the cleavage and formation of α-1,6 glycosidic bonds.
Pathway Questions 4. What are examples of glycogen metabolism in real life? The performance of elite endurance athletes can benefit from a diet regimen of carbohydrate "loading" prior to competition. Carbohydrate loading regimens can result in a build-up of stored muscle glycogen that is sometimes higher than what can be obtained by simply following a high carbohydrate diet. Key is to deplete glycogen before carbo loading to get 2x glycogen level.
Function of Glycogen Phosphorylase Glycogen degradation is initiated by glycogen phosphorylase, a homodimer that catalyzes a phosphorolysis cleavage reaction of the α-1,4 glycosidic bond at the nonreducing ends of the glycogen molecule. Inorganic phosphate (Pi) attacks the glycosidic oxygen using an acid catalysis mechanism that releases glucose-1P as the product. Although the standard free energy change for this phosphorylysis reaction is positive (ΔGº' = +3.1 kJ/mol), making the reaction unfavorable, the actual change in free energy is favorable (ΔG' = -6 kJ/mol) due to the high concentration of Pi relative to glucose-1P inside the cell (ratio of close to 100).
Structure of Glycogen Phosphorylase Exists as a dimer and has binding sites for glycogen and catalytic sites that contain pyridoxal phosphate(derived from vitamin B6). The critical Pi substrate is bound to the active site by interactions with pyridoxal phosphate and active site amino acids.
Function of Phosphoglucomutase The the next reaction in the glycogen degradation pathway is the conversion of glucose-1P to glucose-6P by the enzyme phosphoglucomutase. The enzyme first donates a phosphate group to the substrate to generate an intermediate bisphosphate compound, and then the bisphosphate compound is dephosphorylated to regenerate the phosphoenzyme and release the product. Have you seen this type of reaction before, was it in glycolysis?
Glycogen Debranching Enzyme The glycogen debranching enzyme (also called α-1,6-glucosidase) recognizes the partially degraded branch structure and remodels the substrate in a two step reaction. 1) the debranching enzyme transfers three glucose units to the nearest nonreducing end to generate a new substrate for glycogen phosphorylase. 2) the bifunctional debranching enzyme cleaves the α-1,6 glycosidic bond to release free glucose. Since α-1,6 branch points occur about once every 10 glucose residues in glycogen, complete degradation releases ~90% glucose-1P and 10% glucose molecules. Is there a difference in the amount of energy that can be recovered from glucose-1P and glucose?
Regulation of Glycogen Phosphorylase Activity Activity is regulated by both covalent modification (phosphorylation) and by allosteric control (energy charge). Glycogen phosphorylase is found in cells in two conformations: • active conformation, R form • inactive conformation, T form Phosphorylation of serine 14 (Ser 14) shifts the equilibrium in favor of the active R state. This phosphorylated form of glycogen phosphorylase is called phosphorylase a, and the unphosphorylated form is called phosphorylase b.
Regulation of Glycogen Phosphorylase Activity The enzyme responsible for phosphorylating glycogen phosphorylase b to activate it, is phosphorylase kinase which is a downstream target of glucagon and epinephrine signaling.
Regulation of Glycogen Phosphorylase Activity Insulin stimulates the activity of protein phosphatase-1 (PP-1),leading to inactivation of glycogen phosphorylase. PP-1 is the same enzyme that dephosphorylates PFK-2/FBPase-2, the enzyme responsible for controlling fructose-2,6-BP levels.
Tissue-specific isozymes of glycogen phosphorylase The activity of glycogen phosphorylase can also be controlled by allosteric regulators, which bind to the enzyme and shift the equilibrium. Liver and muscle isozymes of glycogen phosphorylase are allosterically-regulated in different ways, which reflects the unique functions glycogen in these two tissues.
Tissue-specific isozymes of glycogen phosphorylase Liver glycogen phosphorylase a, but not muscle glycogen phosphorylase a is subject to allosteric inhibition by glucose binding which shifts the equilibrium from the R to T state. When liver glycogen phosphorylase a (phosphorylated form) is shifted to the T state, it is a better substrate for dephosphorylation by PP-1 than is the R state. Why does it make sense that muscle glycogen phosphorylase b, but not liver glycogen phosphorylase b, would be allosterically activated by AMP in the absence of hormone signaling?
Glycogen synthase catalyzes glycogen synthesis The addition of glucose units to the nonreducing ends of glycogen by the enzymeglycogen synthase requires the synthesis of an activated form of glucose called uridine diphosphate glucose (UDP-glucose). Glucose-6P is first converted to glucose-1P by phosphoglucomutase, and then the enzyme UDP-glucose pyrophosphorylase catalyzes a reaction involving the attack of a phosphoryl oxygen from glucose-1P on the gamma phosphate of uridine triphosphate (UTP). The rapid hydrolysis of PPi by the abundant cellular enzyme pyrophosphatase results in a highly favorable coupled reaction. Why does rapid conversion of PPi --> 2 Pi result in a more favorable reaction?
Glycogen Synthase Reaction Glycogen synthase transfers the glucose unit of UDP-glucose to the C-4 carbon of the terminal glucose at the nonreducing end of a glycogen chain. The UDP moiety is released and UTP is regenerated in a reaction involving ATP and the enzyme nucleoside diphosphate kinase.
Glycogen Branching Enzyme Once the chain reaches a length of 11 glucose residues, the glycogen branching enzyme transfers seven glucose units from the end of the chain to an internal position using a α-1,6 branchpoint. This new branchpoint must be at least four glucose residues away from the nearest branch. Not the exact reverse of the debranching enzyme’s reaction!
Regulation of Glycogen Synthase Activity The activity of glycogen synthase is also primarily controlled by reversible phosphorylation. The effect of phosphorylation on the activity of glycogen synthase is however the reverse of what we saw with glycogen phosphorylase. Dephosphorylation activates glycogen synthase, whereas, glycogen phosphorylase is activated by phosphorylation. In this case, the active glycogen synthase a form is dephosphorylated and favors the R state, whereas, the inactive glycogen synthase b form is phosphorylated and favors the T state. The “a” form is always the active form; glycogen phosphorylase “a” is phosphorylated, whereas, glycogen synthase “a” is dephosphorylated.
Regulation of Glycogen Synthase Activity Hormone activation of glycogen synthase activity is mediated by insulin, which promotes the activation of glycogen synthase by stimulating PP-1 activity. Epinephrine and glucagon signaling leads to inactivation of glycogen synthase.
Reciprocal regulation of glycogen metabolism Since glycogen phosphorylase and glycogen synthase have opposing effects on glycogen metabolism, it is critical that their activities be reciprocally regulated to avoid futile cycling and to efficiently control glucose-6P concentrations within the cell. Infusion of glucose into mouse liver results in a rapid decrease in glycogen phosphorylase activity within 1 minute, followed by a dramatic increase in glycogen synthase activity by 4 minutes. What is the metabolic logic of glucose inhibition of glycogen phosphorylase activity and activation of glycogen synthase?
Hormone signaling in liver cells insulin signaling on glycogen metabolism in liver cells where it can be seen that glucagon stimulates glucose efflux and insulin stimulates glucose influx through the GLUT2 glucose transporter protein. Net phosphorylation drives glycogen degradation, and net dephosphorylation drives glycogen synthesis.
Glucagon signaling Glucagon signaling in liver cells results in Gsα-mediated stimulation of adenylate cyclase activity leading to the production of the second messenger cyclic AMP. Activation of protein kinase A (PKA) by cAMP triggers two types of phosphorylation circuits in muscle cells; one that stimulates glycogen degradation and a second that inhibits glycogen synthesis.
Insulin signaling Insulin signaling results in dephosphorylation of glycogen metabolizing enzymes and elevated rates of glycogen synthesis. Activation of insulin receptor tyrosine kinase activates the phosphoinositide-3-kinase (PI-3K) pathway which phosphorylates AKT kinase. In turn, Akt phosphorylates glycogen synthase kinase 3 (GSK3) which is inactivated by phosphorylation. Without active GSK3 around to maintain glycogen synthase in the inactive phosphorylated state, the level of active dephosphorylated glycogen synthase increases.
Human glycogen storage diseases A number of human diseases have been identified that affect glycogen metabolism. Disease symptoms in many cases include: liver dysfunction due to excess glycogen fasting-induced hypoglycemia (low blood glucose levels) in the most severe diseases, death at an early age.
Human glycogen storage diseases von Gierke's disease is due to a deficiency in the enzyme glucose-6-phosphatase which causes a build-up of glycogen in the liver because glucose-6P accumulates and activates glycogen synthase. McArdle's disease harbor defects in muscle glycogen phosphorylase. These individuals suffer from exercise-induced muscle pain due to their inability to degrade muscle glycogen.