Download

1 / 28
280 likes | 611 Views
RNA translatsioon. Valgu biosünteesi üldiseloomustus Translatsiooni etapid: initsiatsioon, elongatsioon, terminatsioon Posttranslatsioonilised modifikatsioonid Valgu biosünteesi regulatsioon ja kontroll. Valgu biosünteesi aparatuuri 3 olulisemat komponenti

E N D
Valgu biosünteesi üldiseloomustus • Translatsiooni etapid: initsiatsioon, elongatsioon, terminatsioon • Posttranslatsioonilised modifikatsioonid • Valgu biosünteesi regulatsioon ja kontroll
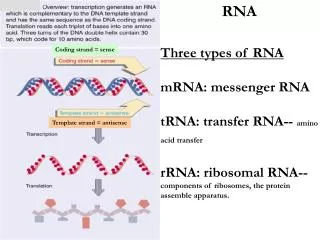
Valgu biosünteesi aparatuuri 3 olulisemat komponenti Aminoatsüül-tRNA aminohapete allikas mRNA- informatsioon valgu sünteesiks Ribosoomid- RNA-st ja valgust koosnevad peptiidahela sünteesi lookused
Valgu biosüntees toimub ribosoomidel Ribosoomid on RNAst ja valkudest koosnevad nukleoproteiidsed kompleksid Ribosoomid koosnevad kahest subühikust E. Coli ribosoomid: Suur subühik 23S rRNA 5S rRNA 49 valku Väike subühik 16S rRNA 33 valku Valgu biosünteesiks on vajalikud mõlemad subühikud
Polüpeptiidi süntees kulgeb C-terminuse suunas Esimene polüpeptiidahelasse lülitatav aminohape on N-terminaalne aminohape ja viimane C-terminaalne aminohape Aminohapped tuuakse riboloomi adaptermolekuli koosseisus Selliseks aktiveeritud aminohappeid ülekandvaks molekuliks on tRNA tRNA on ka vahendiks, mille abil toimub geneetilise koodi (DNA struktuuri) informatsiooni “tõlkimine ehk transleerimine” 20 erineva aminohappe jääkidest koosnevaks valgu struktuuriks tRNA molekul seob oma 3´ terminuses ühe spetsiifilise aminohappe molekuli
Aminohappe aktiveerimine viiakse läbi ensümaatiliselt. Reaktsiooni katalüüsib aminoatsüül-tRNA süntetaas. Süntetaasid peavad ära tundma nii korrektse tRNA kui ka aminohappe Aminohape + ATP aminoatsüül-adenüleet +PPi Aminoatsüül-adenülaat + tRNA aminoatsüül-tRNA + AMP PPi + H2O 2Pi 3´ots, aminohappe sidumine Aminohapped on tRNA-ga seotud estersideme kaudu Antikoodon, mRNA interaktsiooni tsenter
Valgu biosünteesi etapid Esimene etapp- initsiatsioon Initsiatsioonietapis toimub aktiivse sünteesikompleksi moodustumine ribosoomi partiklitest ja mRNAst Initsiatsiooni katalüüsivad spetsiifilised faktorid Initsiatsiooniprotsessis tuntakse ära initsiaatorkoodon AUG Moodustunud initsiatsioonikompleks täidab järgmisi funktsioone fikseerib mRNA korrektselt ribosoomil tekitab struktuurse aluse tRNA-de korrektseks sidumiseks tekitab struktuurse aluse kasvava peptiidi sidumiseks ja väljutamiseks
Valgu biosünteesi etapid Teine etapp- elongatsioon Aminohappe jäägid lisatakse vastavalt mRNA informatsioonile peptiidahela koosseisu Katalüütiline tsenter ribosoomis, kus moodustub peptidüülside, kannab nimetust peptidüültransferaas Ribosoomi liikumine piki mRNA-d, mis on vajalik informatsiooni lugemiseks kannab nimetust translokatsioon Ka elongatsioonil osalevad valgulised faktorid (EF- elongation factor)
Valgu biosünteesi etapid Kolmas etapp- terminatsioon Translatsiooni terminatsiooni signaalina funktsioneerib üks kolmest koodonist (UAA, UAG või UGA), millele ei vasta ühtegi tRNAd Valgu süntees peatub vastava koodonini jõudmisel Spetsiifiliste faktorite (RF- release factor) toimel vabanev kasvav peptiid ribosoomi küljest Ribosoomi kompleks laguneb kaheks subühikuks
Valgu aminohappeline järjestus on määratud mRNA primaarstruktuuri poolt Geneetilist koodi iseloomustab Kood on tripletne- 3 nukleotiidi kodeerib ühe aminohappe Kood ei ole kattuv Kood on pidev (kirjavahemärgidjatühikud puuduvad) Kood on degeneratiivne- ühe aminohappe kodeerimiseks on sageli rohkem kui üks koodon Kood on universaalne- kasutusel ühesugusena peaaegu kõigis organismides. Erandiks on näiteks mitokondrid
Valgu biosünteesi energeetika Aminoatsüül-tRNA sünteesil tekib pürofosfaat, mis hüdrolüüsitakse edasi pürofosfataasi poolt katalüüsitavas reaktsioonis. Ekvivalentne 2 ATP kulutamisega Iga aminohappe jäägi sisenemine peptidüültransferaasi tsentrisse on seotud GTP hüdrolüüsiga- 1 ATP ekvivalent Iga translokatsiooni etapp- seotud GTP hüdrolüüsiga Seega on 1 aminohappe jäägi lülitamine peptiidahelasse seotud vähemalt 4 ATP ekvivalendi kasutamisega Kokku kulutatakse sünteesil 4x30.5=122 kJ/mol Peptiidsideme hüdrolüüs -21 kJ/mol Protsessi pöördumatuse ja täpsuse hind 101 kJ/mol
Posttranslatsioonilised protsessid Valkude pakkumine Valkude keemiline modifitseerimine fosforüleerimine, hüdroksüleerimine glükosüleerimine, lipiidsete konjugaatide moodustumine Valkude primaarstruktuuri moodusumine ei ole sageli piisav bioloogiliselt aktiivse valgumolekuli moodustumiseks
Valkude pakkumine Valgomolekuli ruumilise struktuuri teke sõltub peptiidahela ja kõrvalahelate funktsionaalrühmade interaktsioonidest Lisaks spontaansele, mittekatalüütilisele protsessile osalevad tsaperonid prolüül cis-trans isomeraas Kõrvalahelate interaktsioonid Ioonsed sidemed Hüdrofoobne interaktsioon Dipoolide vaheline interaktsioon Kovalentne side
Biokeemilised valkude modifikatsoonid Spetsiifiline proteolüütiline hüdrolüüs Umbes pooltel valkudel eemaldatakse N-terminaalne aminohape (Met) Paljud ensüümid sünteesitakse inaktiivsete prekursoritena- tsümogeenide kujul. Tsümogeenide aktivatsiooniks on vajalik spetsiifiliste peptiidsidemete hüdrolüüs
Biokeemilised valkude modifikatsoonid Aminohappe jääkide modifikatsioon Fosforüleerimine Fosforüleeritakse sageli seriini, treoniini ja türosiini jääke. Fosforüleerimine on sageli pöörduv Hüdroksüleerimine Proliini ja lüsiini modifitseerimine hüdroksüproliiniks ja hüdroksülüsiiniks
Biokeemilised valkude modifikatsoonid Suhkrute kovalentne sidumine Glükoproteiinide moodustumisel modifitseeritakse kas seriini, treoniini (O seoseline glükosüleerimine) või asparagiini jääke (N-seoseline glükosüleerimine) Prosteetiliste rühmade lisamine Väikeste orgaaniliste molekulide, organometalliliste rühmade või anorgaaniliste molekulide sidumine- heem, FAD, biotiin
Valkude suunamine raku organellidesse ja rakust välja Valkude sünteesilookusest tuleb valgud transportida sinna, kus nende funktsioon on vajalik Prokarüoodid- osa valke tuleb rakust väljutada Eukarüoodid- ribosoomid paiknevad tsütoplasmas. Loetud arv valke sünteesitakse ka mitokondris. Ülejäänud organellidesse ja ka suur osa mitokondris paiknevaid valke tuleb sinna transportida. Transportsignaali mehhanism- üldiselt sisaldavad transporditavad valgud teatud signaaljärjestust, mis tüüpiliselt on 15-40 aminohapet molekuli N terminuses Signaaljärjestus eemaldatakse hüdrolüütiliselt peale transporti korrektsesse lookusesse Tuuma transpordi signaal paikneb tavaliselt molekuli keskel ja seda ei hüdrolüüsita
Valgu taseme regulatsioon Aktiivse valgu hulk sõltub ühelt poolt sünteesi kiirusest ja teiselt poolt degradatsioonist. Valkude degradatsioon Valkude degradatsioon on reguleeritud, mida näitavad erinevused valkumolekulide elueas Valk poolestusaeg RNA polümeraas I (roti maks) 1.5 min Tsütokroom C (roti maks) 150 min Hemoglobiin (inimese) 100 päeva Kahjustunud või oma ülesande täitnud valgumolekulide suunamiseks degradatsiooni ratta on olemas spetsiifilised mehhanismid
Valkude degradatsioon Ubikitiini rada Ubikitiini rada on degradeerimiseke suunatud valkude spetsiifiline märgistamise mehhanism eukarüootsetes rakkudes Ubikitiin- väike valk 76 aminohappe jääki Kõrgelt konserveerunud, pärmi ja inimese ubikitiini erinevus on 3 aminohappe jääki Ubikitiin seotakse kovalentselt märklaud valgu lüsiini jääkidega Tavaliselt on mitu ubikitiini molekuli seotud ühe valgu molekuliga Ubikitiiniga märgistatud valgud degradeeritakse proteolüütiliselt
Valgu biosünteesi regulatsioon Valke kodeerivate geenide arv bakter 4000 pärm 6000 Inimene 100,000 Vaid murdosa nendest tuhandetest valkudest on ühes rakus konkreetsetes tingimustes aktiivselt sünteesitav Kuidas toimub sünteesi regulatsioon? Kõige olulisem on kontroll transkriptsiooni initsiatsiooni tasemel transkriptsioon post-transkriptsiooniline protsessing mRNA degradatsioon translatsioon post-translatsiooniline protsessing
Geeni ekspressiooni regulatsioon Geenide ekspressiooni reguleeritakse kolmel fundamentaalselt erineval printsiibil: Konstitutiivselt ekspresseeritud geenid Transkripti toodetakse pidevalt, mistõttu produtseeritakse teatud konstantsel tasemel produkti (valku) Indutseeritavad geenid- teatud faktorite toimel aktiveeritavad geenid Represseeritavad geenid- teatud faktorite toimel deaktiveeritavad geenid Viimase kahe klassi transkripti hulk ei ole konstantne vaid muutub keskkonna tingimustest, kasvufaasist, rakutsüklist jm. sõltuvalt
Geeni ekspressiooni regulatsioon Sageli paiknevad seotud funktsiooniga valkude geenid prokarüootsetes rakkudes genoomis klastritena Sellise operonstruktuuri moodustavad Struktuurgeenid, mida transkribeeritakse ja transleeritakse Promootorregioon- vastutab RNA polümeraasi sidumise eest Induktorite sidumisregioon Repressorite sidumisregioon- operaatorid Kõik struktuurgeenid on operonis reguleeritud koos ühise mehhanismiga
Geeni ekspressiooni regulatsioon RNA polümeraas RNA polümeraas on transkriptsiooni protsessi kõige olulisem komponent RNA polümeraas initseerib transkriptsiooni seondudes promootorregiooniga DNA struktuuris Promootorregiooni struktuursed iseärasused määravad ära RNA polümeraasi seondumise efektiivsuse Indutseeritavate või represseeritavate geenide korral toimib täiendav kontroll transkriptsiooni initsiatsiooni üle. Harilikult toimib see kontroll regulatoorsete valkude abil, mis seonduvad DNA-ga. Geeni ekspressiooni regulatsioon eukarüootides on tunduvalt keerukam Promootorregiooni regulatoorsed elemendid on kompleksed RNA polümeraase on 3
Geeni ekspressiooni regulatsioon Mehhanitsistlikult võib reguleeritavad protsessid jaotada positiivse ja negatiivse regulatsiooniga protsessideks Positiivne regulatsioon Transkriptsioon toimub molekulaarse signaali olemasolu korral Negatiivne regulatsioon Transkriptsioon blokeeritakse molekulaarse signaali korral Regulatsioon toimib tüüpiliselt üle regulatoorsete valkude DNA-ga seostumise muutuse
Geeni ekspressiooni regulatsioon Regulatoorseid valke iseloomustavad teatud sageli esinevad diskreetsed DNA-ga seonduvad domeenid 20...100 aminohappe jääki Domeenide struktuur vastab DNA kaksikheeliksi struktuurile Moodustavad suhteliselt tugeva kompleksi spetsiifilist järjestust omava DNA fragmendiga, rakendades spetsiifilisi mittekovalentseid interaktsioone, muuhulgas vesiniksidemeid, mida kaksikheeliks võimaldab
Geeni ekspressiooni regulatsioon Regulatoorsete valkude klassid Heeliks-pööre-heeliks (helix-turn-helix) Ca 20 aminohappe pikkune domeen, koosneb kahest heeliksist ja neid ühendavast pöördest
Geeni ekspressiooni regulatsioon Regulatoorsete valkude klassid Zn-finger motiiv Eukarüootse päritoluga regulatoorsed valgud, sisaldavad Zn2+ iooni, mis on koordineeritud Cys ja His jääkide poolt
Geeni ekspressiooni regulatsioon Regulatoorsete valkude klassid Leucine zipper- leutsiini tõmblukk Heeliksi regioon, kus leutsini jääk on igas 7-ndas positsioonis, mis paiknevad heeliksi ühel küljel. Osaleb hüdrofoobsetes interaktsioonides