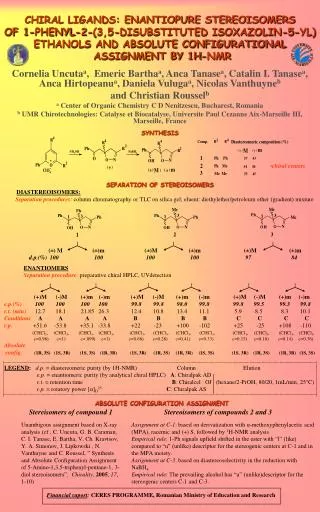
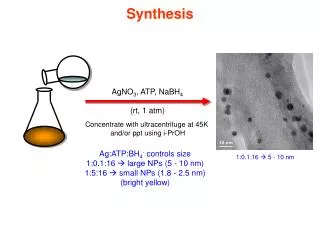
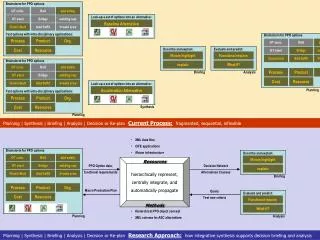
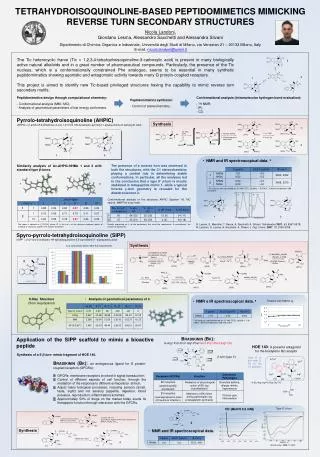
Download

1 / 28
280 likes | 335 Views
Synthesis. Genetics and natural selection combine. Norton and Hardy. Statistical calculations begin to show that an advantageous gene can spread quite quickly through a population– even if it only provides a small advantage.

E N D
Synthesis Genetics and natural selection combine
Norton and Hardy • Statistical calculations begin to show that an advantageous gene can spread quite quickly through a population– even if it only provides a small advantage. • Hardy-Weinberg equilibrium expresses how the proportion of various genotypes (in a population ‘at equlibrium’ i.e. with no net selection acting on it) is determined by the frequencies of the alternative alleles for a particular gene: Where p is the proportion of alleles of form A, and q is the proportion of alleles of form a, the proportion of genotypes in the next generation (given random mating) is: • p2 (of type AA): 2pq (of type Aa): q2 (of type aa).
Calculating genetic change • Consider Norton’s table (p. 184). • Let’s think about the numbers here and why they vary as they do. • First think of the difference that the different strengths of the selection effect makes. • Then think of the difference that dominance vs. recessiveness of the new gene makes. • Take-home: Natural selection can shift the frequencies of genes in a population fairly quickly.
Ronald Fisher • Fisher was a statistician with a serious interest in evolution. • His statistical analysis of gene selection showed that evolution could proceed gradually using small mutations instead of suddenly forming new species by a single mutational ‘jump’.
The Genetical Theory of Natural Selection • Published in 1930, this book explained in detail how gradual evolution could be accomplished under Mendelian genetics. • Chance survival or loss of genes is important only for very small populations. • Mutations were sufficiently rare that only with the help of selection could they be expected to add up and create substantial evolutionary change in a population. • Even directed mutations can’t resist the impact of selection forces: orthogenesis is ruled out.
The upshot • Contra de Vries and Bateson (and the early Thomas Morgan) the only defensible theory of evolution, given Mendelian genetics, is evolution by natural selection. • British biologist J.B.S. Haldane reached the same conclusions independently, on the basis of his own mathematical analysis of genetics and natural selection. • Statistical analysis of the changing population showed that the dark form of the peppered moth (Biston betularia) had enjoyed a 30% selective advantage over the normal form. The intensity of selection in wild populations is sometimes quite high!
Polymorphism • Populations don’t always tend to ‘fix’ one allele at each position. • Sometimes the heterozygotes (those with both forms of the allele) actually have an advantage– in this case, selection will tend to preserve both alleles, at proportions that balance the disadvantages of each homozygotic form and the advantage of the heterozygotes. • Sometimes combinations of genes contribute to selective advantages and disadvantages– this can also lead selection to maintain polymorphism in a population.
William Castle • Ovary transplants (once more, inheritance of acquired characteristics fails the test). • Genes interact (hooded gene in rats interacts with genes biasing the pattern towards all-black and all-white coats). • So genotype does not map ‘1 to 1’ into phenotype. • This pleiotropy unites each organism into a distinct individual through the subtle effects of the particular gene combinations it has. • It also ensures that there is a substantial pool of genetic variation for selection to operate on.
Sewall Wright • Student of Castle. • Involved in these experiments. • Also mathematically gifted, like Fisher and Haldane. • But involved in agricultural breeding work as well.
A strategy for breeding • Wright was interested in exploring rare combinations of genes for their potential in breeding new and desirable traits. • His strategy was to use inbreeding to concentrate particular combinations of pure (homozygotic) traits. • Then crossbreeding the best of these animals could produce desirable combinations of traits that weren’t visible in the original population because the combinations were so unlikely under random mating. • This kind of breeding has contributed to new crops and new food animals (such as dairy cattle) that are far more productive than their ancestors were.
In nature • The same could occur in nature, in a large population made up of mostly-isolated subpopulations. • The variation and different combinations of genes that arise in different subpopulations would allow for the rapid accumulation of advantages in one population or another, which could then spread throughout the larger population over time. • Here Wright differed from Fisher and Haldane, who thought of variation at a more individual level.
Reconciliation • This work offered a reconcilliation between the geneticists and the naturalists. • Naturalists focused on continuous variation in natural populations and how selection would act on that variation over time. • Geneticists focused on discrete (even dramatic) differences and their origins as ‘mutations’. • But small mutations, as Fisher, Haldane and Wright showed, could spread through a population by natural selection, producing the continuous variation and slow change the naturalists believed in using the mechanisms that the geneticists accepted.
Isolation and speciation • Restrictions on interbreeding lead to local ‘races’ or subspecies. • The line between subspecies and species was and remains a vague one. • Darwin distinguished evolution as change in a single lineage over time and evolution in the ‘branching’ sense, in which a single lineage separates into two (or more). • This, he thought, would normally require some degree of isolation.
Wagner • Distinct but close species are often separated by a geographical barrier (mountain range, or even a ridge separating one valley from another, or a river, or…) • Believed this was the key to speciation. • Tried to make this into a general theory of evolution separate from natural selection. • But not all isolation is geographical– ecological patterns could also separate two lineages.
Gulick • Separation by degrees in land snails. • A big lesson: the same phenomenon here, laid out spatially, as we encounter with all species when we think in terms of long periods of time: • Each small unit of change is not enough to make a new species– but as we follow from one group to the next, we get new species and eventually even new genera.
Sumner and subspecies of mice • Genetic/ breeding studies on different subspecies (lighter and darker) of peromyscus mice showed the standard pattern of ‘Mendelian segregation’: In the first generation, colours were intermediate, but the full range reappeared in the second generation, with intermediates indicating several genes were involved in the differences. • The differences were not driven by climate– the shift from one to the other form took place in as little as 10 miles. • Environment (light sand beaches vs. darker forest floors) provided a natural explanation of the differences, in terms of natural selection.
Dobzhansky: the synthesis applied to natural field studies • Theodosius Dobzhansky left Russia to work with Thomas Morgan at Columbia, and then followed Morgan to Stanford. • He was the first biologist to integrate detailed field studies of variation in species with genetics and natural selection. • One example of natural selection he studied was the development of pesticide resistance in scale insects. • Resistance had originated in small areas, and then spread to other areas– just as if it were due to a mutation in one population that then spread as the insects that carried it survived and spread while others died. • For Dobzhansky, evolution just is change in the frequencies of genes in a population over time.
The idea of a species • When they inhabit the same territory, species are clearly, sharply distinct. • But when we trace them across different regions, we find differences– and the differences tend to increase the further we go. • In fact, there are things called ring species, in which a group of interbreeding populations stretch out across a ring of territory, but the populations at opposite ends of the ring, which overlap territorially, are distinct: They don’t interbreed at all. • Northern gulls are one example.
So what is a species? • Aristotle thought in terms of essences: A species was all the living things that shared an essence. • For Aristotle, these essences are preserved (passed on unchanged) in reproduction– so they can’t change at all. • Thinking in this way makes the ‘type’ more real and permanent– it’s because they have the type’s unchanging essence that individuals belong to a particular species. • But when we look at the world, what we see first isn’t types– it’s individual organisms, all of which vary to one degree or another.
Populations • Instead of thinking in terms of types, biologists like Darwin and Wallace began to think in terms of populations– groups of very similar organisms that reproduce, sharing their genes through the generations. • This makes the individual organism the starting point, instead of the type. • And it raises questions about essences: Is there really some basic set of characteristics that constitute a type for each species? Ring species make this question particularly hard to answer.
Mayr: The synthesis applied to taxonomy • Mayr, like Dobzhansky, came to think of species in a more population-based way. • What unites a population over time is interbreeding. • So Mayr followed Dobzhansky in setting the line between species at the point where two populations would not interbreed if they shared the same territory. • This can be due to sterility, behavioural differences, differences in mating preferences, or a combination of these. • This is often called the biological species concept. • Mayr was able to trace the development of species of tropical birds using this criterion.
Lack • Lack applied the same ideas to Darwin’s finches on the Galapagos. • Studying them in greater detail, Lack showed that there were 14 species of finch on the Galapagos. • He also established how they had spread from island to island and gradually developed into distinct species that sometimes now lived on the same island. • Lack also showed that the differences in beak shape in the finches were adaptations to different diets.
Simpson: the synthesis applied to paleontology • Paleontology was the last field of biology to adopt the synthesis. • Paleontologists like Cope had long focused on ‘evolutionary trends’ which they could see in the fossil record. • Thinking in terms of trends made natural selection less convincing to them– orthogenesis remained popular despite the arguments of Fisher, Haldane and Wright. • Simpson’s mentor, Osborn, was student of Cope’s and loyal to the traditional view.
Tempo and Mode in Evolution • Simpson brought the synthesis to bear on paleontological evidence in this book (1944). • He used fossils to estimate the rate of evolutionary change, and compared these rates to those that had been observed in nature by people like Dobzhansky. • Change proceeded at varying rates in the fossil record, from quite quickly to very slowly. • Simpson also emphasized the ‘bushy’ quality of evolution– instead of following a single lineage to trace the history of a present-day group, he tried to look at the history of the whole family of groups, living and extinct, that derived from a common ancestor.
Natural selection • The pattern was not one of linear trends– evolution had moved at different rates, and in different directions from time to time and place to place. • This made perfect sense from the point of view of natural selection. • Naturalists, taxonomists and paleontologists were now all on board– the modern synthesis had been fully integrated into the main fields of biology.