Download
1 / 12
120 likes | 191 Views
Learn about glycine titration and isoelectric point through its forms and reaction at different pH values. Fully explained with diagrams and calculations.

E N D
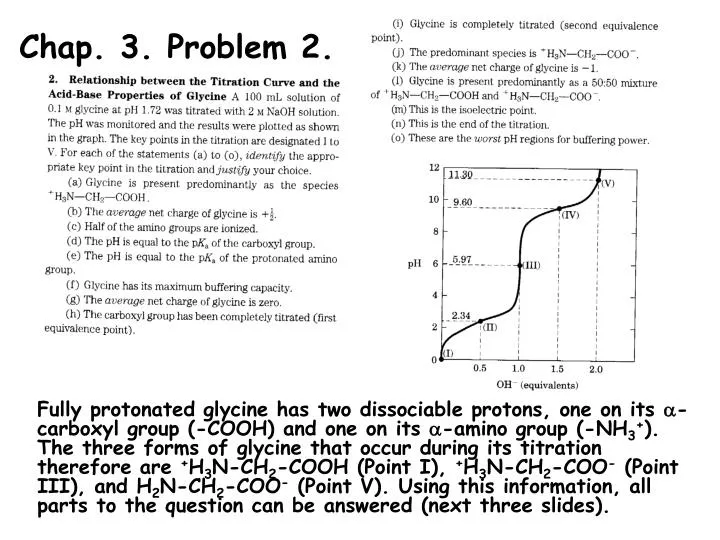
Chap. 3. Problem 2. Fully protonated glycine has two dissociable protons, one on its -carboxyl group (-COOH) and one on its -amino group (-NH3+). The three forms of glycine that occur during its titration therefore are +H3N-CH2-COOH (Point I), +H3N-CH2-COO- (Point III), and H2N-CH2-COO- (Point V). Using this information, all parts to the question can be answered (next three slides).
Chap. 3. Problem 2. (continued) Part (a). The fully protonated species of glycine (+H3N-CH2-COOH) occurs at the beginning of the titration, i.e., Point I. Part (b). 50% of the -COOH protons are titrated to -COO- at the pK1 of glycine (Point II, pH 2.34). At this point, half of the molecules have a net +1 charge (+H3N-CH2-COOH) and half have a 0 charge (+H3N-CH2-COO-). Thus the average net charge of glycine is +1/2 at Point II. Part (c). Half of the -amino groups of glycine molecules are ionized at its pK2 (Point IV, pH 9.6). Part (d). From the HH equation, a 1/1 ratio of the conjugate base/conjugate acid forms of a weak acid occur when pH = pKa. Thus, the pH of the glycine solution is equal to the pKa of the -carboxyl group at Point I where, 50% of the -COOH groups have been titrated. Part (e). Point IV. See the explanation in the answer to Part (d). Part (f). The optimum buffering pHs for a solution of glycine occur at the two plateaus in the titration curve (Points II & IV). This is where the concentrations of the conjugate base and conjugate acid forms undergoing titration are the highest.
Chap. 3. Problem 2. (continued) Part (g). The average net charge of glycine is zero at its isoelectric point. This occurs after the -COOH group of the amino acid has been titrated and the most prevalent species in solution has the structure, +H3N-CH2-COO-. This is Point III, which occurs at a pH halfway between pK1 and pK2 pI = (pK1 + pK2)/2 = (2.34 + 9.60)/2 = 5.97. Part (h). The first equivalence point in the titration occurs when the -COOH group has been completely titrated, i.e., Point III. Part (i). The second equivalence point in the titration occurs when the -NH3+ group has been completely titrated, i.e., Point V. Part (j). The species, +H3N-CH2-COO-, which has a net charge of zero occurs at the pI. This species predominates at Point III, which has a pH of 5.97. Part (k). The average net charge of glycine occurs at Point V, where both dissociable groups have been completely titrated and the structure of the molecule is H2N-CH2-COO-. Part (l). Glycine occurs with a 50/50 mixture of +H3N-CH2-COOH and +H3N-CH2-COO- when the -COOH group has been 50% titrated. This is at the midpoint of the first plateau in the titration curve, where pH = pK1.
Chap. 3. Problem 2. (continued) Part (m). The isoelectric point occurs at Point III. (See answers (g) and (j). Part (n). Glycine has two dissociable groups, so the end of the titration occurs when 2.0 equivalents of strong base have been added. This corresponds to Point V on the curve. Part (o). The worst pH regions for glycine to serve as a buffer occur at the points furthest from the two plateaus, pH = pK1 and pH = pK2. These regions occur at Points I, III, and V.
Chap. 3. Problem 4. Part (a). The structures, equilibrium equations, and pKas for the three ionization reactions of histidine are shown in the diagram. The net charge on histidine in each ionization state is (1) +2; (2) +1; (3) 0; and (4) -1. Part (b). The structures are the same as in Part a because each pH listed flanks the pKas of histidine’s three dissociable protons. Part (c). The net charge of the histidine species at each of these pHs is shown in the figure. Based on the net charge, histidine will migrate towards the (1) cathode; (2) cathode; (3) not migrate; and (4) anode when placed in an electric field.
Chap. 3. Problem 8. The average Mr of an amino acid in a protein is 128. After correcting for the loss of water (Mr 18) that occurs in peptide bond formation the average Mr becomes 110. Based on this value, the size of the protein is approximately 682 aa x 110 aa-1 = 75,020 = 75,000 = Mr
Chap. 3. Problem 10. Both types of electrophoresis used in this question separate polypeptides on the basis of molecular weight. The second procedure also breaks intra- and intermolecular disulfide bonds that may be present. Because the 160 and 60 kDa polypeptides are present under both conditions, the data suggest that these two proteins are not attached to others by disulfide bonds. Because the 180 kDa protein obtained in the first procedure disappears and a new band at 90 kDa appears, it suggests that the 180 kDa protein may actually consist of two 90 kDa subunits. Thus the most likely subunit composition of this 400 kDa protein is (160, 902, 60). The two 90 kDa subunits are linked by an intermolecular disulfide bond(s).
Chap. 3. Problem 13. Like amino acids, peptides and proteins have isoelectric points at which the sum of all charged residues on the protein is zero. If a protein has a relatively large number of basic residues (His, Arg, Lys) then the pI of the protein will be high. Conversely, if it has a relatively large number of acidic residues (Asp, Glu), then the protein will have a low pI. Histones have high pI values because they have large numbers of His, Arg, and Lys residues. Because the side-chains of these residues are partially (His) or fully (Arg & Lys) positively charged at neutral pH, histones bind strongly to the negatively charged phosphate groups of DNA via ionic interactions.
Chap. 3. Problem 15. Part (a). The specific activity (S.A.) of an enzyme sample is defined as the number of units of activity divided by the total mg of protein in the sample (U/mg). Using this formula, the S.A. obtained after each step of the purification listed in the table is Step 1) 200; 2) 600; 3) 250; 4) 4,000; 5) 15,000; and 6) 15,000 U/mg. Part (b). The most effective step in the purification is the one that results in the greatest increase in S.A. of the enzyme. This corresponds to Step 4 (ion-exchange chromatography) wherein the S.A. increased by 16-fold (4,000/250). Part (c). By a similar logic to that used in Part (b), Step 3 (pH precipitation) is the least effective step of the procedure. Two-thirds of the total activity of the protein actually was lost in this step. Part (d). The enzyme is pure after Step 6 based on the fact that the S.A. did not increase between Steps 5 and 6. SDS PAGE could be used to assess the final purity of the enzyme.
Chap. 3. Problem 18. Part (a). The amino acid composition of the peptide is obtained using this procedure. Thus we can write the empirical composition of the peptide as (Gly2, Leu1, Phe1, Tyr1)n, where n indicates that these residues theoretically could be present in two or more copies. Part (b). FDNB is used to determine the N-terminal amino acid of a peptide or polypeptide. Since no other Tyr residue was found following complete hydrolysis of the FDNB-labeled peptide, this indicates that n = 1. At this stage of the analyses, the sequence of the peptide can be written as Tyr-(Gly2, Leu1, Phe1), where the last four residues could occur in any order. Part (c). The enzyme chymotrypsin cleaves polypeptides on the carboxyl side of aromatic residues (Phe, Trp, and Tyr). Since three products (Tyr, Leu, and the tripeptide [Phe1, Gly2]) were obtained by enzymatic cleavage, the data indicate that Phe must be internal to the peptide sequence and not at the C-terminus. Thus Leu must be at the C-terminus. The only sequence consistent with the data therefore is Tyr-Gly-Gly-Phe-Leu.
Chap. 3. Problem 21. The conventions used in writing consensus sequences in proteins are explained in Box 3-2. Also refer to Table 3-1 for amino acid symbols. Part (a). The invariant residues in this sequence are Tyr (Y) 1, Phe (F) 7, and Arg (R) 9. Part (b). Positively charged amino acids must be present at positions 4 and 9. Lys (K) is more common at position 4, whereas Arg (R) is invariant at position 9. Part (c). Acidic amino acids with negatively charged side-chains must be present at positions 5 and 10. Glu (E) predominates at both of these positions. Part (d). As indicated by the x in the sequence, position 2 can be any amino acid. However, Ser (S), appears most often there.
Part (a). Peptide 2 has the highest proportion of negatively charged to positively charged side-chains (7 to 1, out of 18 total residues). Thus, it will migrate the slowest through an anion exchange column which has positively charged groups bound to the resin. Part (b). Peptide 1 has the highest proportion of positively charged to negatively charged side-chains (11 to 1, out of 44 total residues). Thus, it will migrate the slowest through an cation exchange column which has negatively charged groups bound to the resin. Part (c). A gel filtration column separates proteins on the basis of size. Since peptide 2 is the smallest, it will move through a gel filtration column at the slowest rate. Part (d). Peptide 3 has the sequence G-x(2)-G-x-G-K-S which corresponds to the logo. Chap. 3. Problem 22.