Download
1 / 1
10 likes | 74 Views
Explore how an unidentified transcription factor specific to CD4 lineage may direct lineage commitment by binding the Cd4 promoter site P3, impacting T cell development. Use Yeast one-hybrid system to identify this factor.

E N D
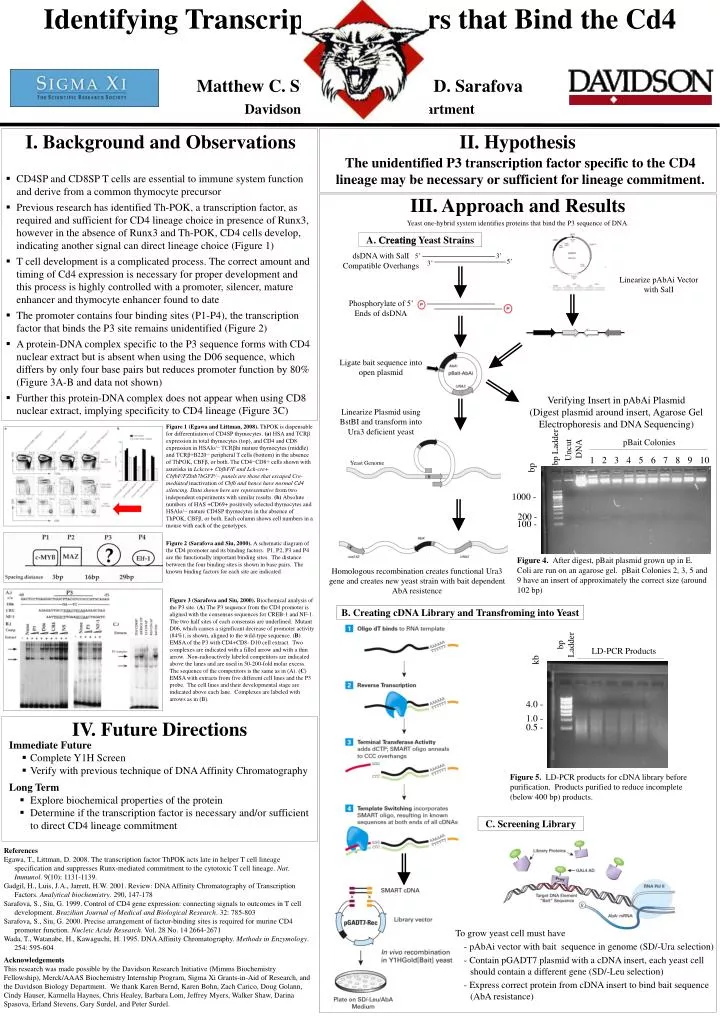
Identifying Transcription Factors that Bind the Cd4 Promoter Matthew C. Surdel and Sophia D. Sarafova Davidson College Biology Department • I. Background and Observations • CD4SP and CD8SP T cells are essential to immune system function and derive from a common thymocyte precursor • Previous research has identified Th-POK, a transcription factor, as required and sufficient for CD4 lineage choice in presence of Runx3, however in the absence of Runx3 and Th-POK, CD4 cells develop, indicating another signal can direct lineage choice (Figure 1) • T cell development is a complicated process. The correct amount and timing of Cd4 expression is necessary for proper development and this process is highly controlled with a promoter, silencer, mature enhancer and thymocyte enhancer found to date • The promoter contains four binding sites (P1-P4), the transcription factor that binds the P3 site remains unidentified (Figure 2) • A protein-DNA complex specific to the P3 sequence forms with CD4 nuclear extract but is absent when using the D06 sequence, which differs by only four base pairs but reduces promoter function by 80% (Figure 3A-B and data not shown) • Further this protein-DNA complex does not appear when using CD8 nuclear extract, implying specificity to CD4 lineage (Figure 3C) II. Hypothesis The unidentified P3 transcription factor specific to the CD4 lineage may be necessary or sufficient for lineage commitment. III. Approach and Results Yeast one-hybrid system identifies proteins that bind the P3 sequence of DNA. A. Creating Yeast Strains dsDNA with SalI Compatible Overhangs 5’ 3’ 5’ 3’ Linearize pAbAi Vector with SalI Phosphorylate of 5’ Ends of dsDNA Ligate bait sequence into open plasmid Verifying Insert in pAbAi Plasmid (Digest plasmid around insert, Agarose Gel Electrophoresis and DNA Sequencing) Linearize Plasmid using BstBI and transform into Ura3 deficient yeast Figure 1 (Egawa and Littman, 2008). ThPOK is dispensable for differentiation of CD4SP thymocytes. (a) HSA and TCRβ expression in total thymocytes (top), and CD4 and CD8 expression in HSAlo/− TCRβhi mature thymocytes (middle) and TCRβ+B220− peripheral T cells (bottom) in the absence of ThPOK, CBFβ, or both. The CD4−CD8+ cells shown with asterisks in Lckcre+ CbfbF/F and Lck-cre+ CbfbF/FZbtb7bGFP/− panels are those that escaped Cre-mediated inactivation of Cbfb and hence have normal Cd4 silencing. Data shown here are representative from two independent experiments with similar results. (b) Absolute numbers of HAS +CD69+ positively selected thymocytes and HSAlo/− mature CD4SP thymocytes in the absence of ThPOK, CBFβ, or both. Each column shows cell numbers in a mouse with each of the genotypes. pBait Colonies Uncut DNA bp Ladder 1 2 3 4 5 6 7 8 9 10 bp 1000 - 200 - 100 - Figure 2 (Sarafova and Siu, 2000). A schematic diagram of the CD4 promoter and its binding factors. P1, P2, P3 and P4 are the functionally important binding sites. The distance between the four binding sites is shown in base pairs. The known binding factors for each site are indicated. Yeast Genome Figure 4. After digest, pBait plasmid grown up in E. Coli are run on an agarose gel. pBait Colonies 2, 3, 5 and 9 have an insert of approximately the correct size (around 102 bp) Homologous recombination creates functional Ura3 gene and creates new yeast strain with bait dependent AbA resistence Figure 3 (Sarafova and Siu, 2000). Biochemical analysis of the P3 site. (A) The P3 sequence from the CD4 promoter is aligned with the consensus sequences for CREB-1 and NF-1. The two half sites of each consensus are underlined. Mutant D06, which causes a significant decrease of promoter activity (84%), is shown, aligned to the wild-type sequence. (B) EMSA of the P3 with CD4+CD8- D10 cell extract. Two complexes are indicated with a filled arrow and with a thin arrow. Non-radioactively labeled competitors are indicated above the lanes and are used in 50-200-fold molar excess. The sequence of the competitors is the same as in (A). (C) EMSA with extracts from five different cell lines and the P3 probe. The cell lines and their developmental stage are indicated above each lane. Complexes are labeled with arrows as in (B). B. Creating cDNA Library and Transfroming into Yeast bp Ladder LD-PCR Products kb 4.0 - 1.0 - IV. Future Directions 0.5 - • Immediate Future • Complete Y1H Screen • Verify with previous technique of DNA Affinity Chromatography Figure 5. LD-PCR products for cDNA library before purification. Products purified to reduce incomplete (below 400 bp) products. • Long Term • Explore biochemical properties of the protein • Determine if the transcription factor is necessary and/or sufficient to direct CD4 lineage commitment C. Screening Library References Egawa, T., Littman, D. 2008. The transcription factor ThPOK acts late in helper T cell lineage specification and suppresses Runx-mediated commitment to the cytotoxic T cell lineage. Nat. Immunol. 9(10): 1131-1139. Gadgil, H., Luis, J.A., Jarrett, H.W. 2001. Review: DNA Affinity Chromatography of Transcription Factors. Analytical biochemistry. 290, 147-178 Sarafova, S., Siu, G. 1999. Control of CD4 gene expression: connecting signals to outcomes in T cell development. Brazilian Journal of Medical and Biological Research. 32: 785-803 Sarafova, S., Siu, G. 2000. Precise arrangement of factor-binding sites is required for murine CD4 promoter function. Nucleic Acids Research. Vol. 28 No. 14 2664-2671 Wada, T., Watanabe, H., Kawaguchi, H. 1995. DNA Affinity Chromatography. Methods in Enzymology. 254: 595-604 • To grow yeast cell must have • - pAbAi vector with bait sequence in genome (SD/-Ura selection) • - Contain pGADT7 plasmid with a cDNA insert, each yeast cell should contain a different gene (SD/-Leu selection) • - Express correct protein from cDNA insert to bind bait sequence (AbA resistance) Acknowledgements This research was made possible by the Davidson Research Initiative (Mimms Biochemistry Fellowship), Merck/AAAS Biochemistry Internship Program, Sigma Xi Grants-in-Aid of Research, and the Davidson Biology Department. We thank Karen Bernd, Karen Bohn, Zach Carico, Doug Golann, Cindy Hauser, Karmella Haynes, Chris Healey, Barbara Lom, Jeffrey Myers, Walker Shaw, Darina Spasova, Erland Stevens, Gary Surdel, and Peter Surdel.