Download
1 / 1
10 likes | 91 Views
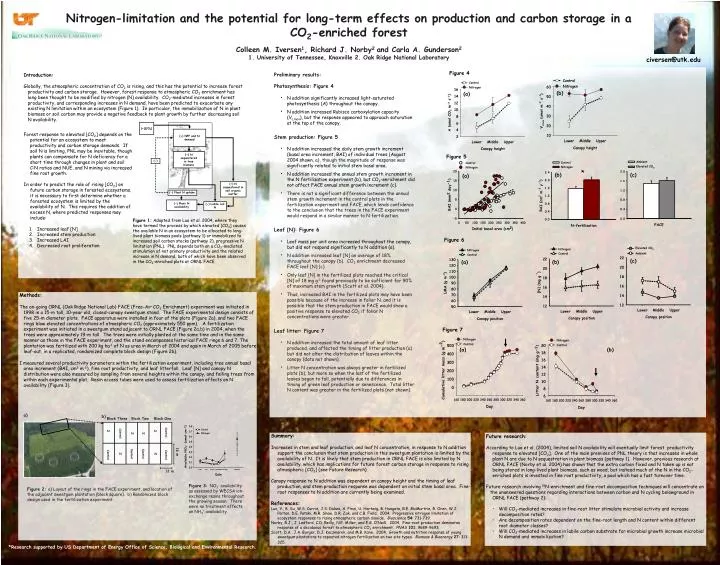
Figure 4. (b). (a). (+)[CO 2 ]. (+) NPP and N demand. (+) N sequestered in tree biomass . Figure 5. (-). *. (+) N sequestered in soil organic matter . (b). (c). (a). (-) Plant N uptake. (-) Plant N availability. (-) Labile soil N. Figure 6. (c). (a). (b). Figure 7. (a). (b).

E N D
Figure 4 (b) (a) (+)[CO2] (+) NPP and N demand (+) N sequestered in tree biomass Figure 5 (-) * (+) N sequestered in soil organic matter (b) (c) (a) (-) Plant N uptake (-) Plant N availability (-) Labile soil N Figure 6 (c) (a) (b) Figure 7 (a) (b) a) b) Block Three Block Two Block One Figure 3: NO3- availability as assessed by WECSA ion-exchange resins throughout the growing season. There were no treatment effects on NH4+ availability. N N Control Control Figure 2: a) Layout of the rings in the FACE experiment, and location of the adjacent sweetgum plantation (black square). b) Randomized block design used in the fertilization experiment. N N 16 m Control Control Control Control N N 12 m Nitrogen-limitation and the potential for long-term effects on production and carbon storage in a CO2-enriched forest Colleen M. Iversen1, Richard J. Norby2 and Carla A. Gunderson2 1. University of Tennessee, Knoxville 2. Oak Ridge National Laboratory civersen@utk.edu • Preliminary results: • Photosynthesis: Figure 4 • N addition significantly increased light-saturated photosynthesis (A) throughout the canopy. • N addition increased Rubisco carboxylation capacity (Vc,max), but the response appeared to approach saturation at the top of the canopy. • Stem production: Figure 5 • N addition increased the daily stem growth increment (basal area increment, BAI) of individual trees (August 2004 shown, a), though the magnitude of response was significantly related to initial stem basal area. • N addition increased the annual stem growth increment in the N fertilization experiment (b), but CO2-enrichment did not affect FACE annual stem growth increment (c). • There is not a significant difference between the annual stem growth increment in the control plots in the fertilization experiment and FACE, which lends confidence to the conclusion that the trees in the FACE experiment would respond in a similar manner to N fertilization. • Leaf [N]: Figure 6 • Leaf mass per unit area increased throughout the canopy, but did not respond significantly to N addition (a). • N addition increased leaf [N] an average of 18% throughout the canopy (b). CO2 enrichment decreased FACE leaf [N] (c) • Only leaf [N] in the fertilized plots reached the critical [N] of 18 mg g-1 found previously to be sufficient for 90% of maximum stem growth(Scott et al. 2004). • Thus, increased BAI in the fertilized plots may have been possible because of the increase in foliar N, and it is possible that the stem production in FACE would show a positive response to elevated CO2 if foliar N concentrations were greater. • Leaf litter: Figure 7 • N addition increased the total amount of leaf litter produced, and affected the timing of litter production (a) but did not alter the distribution of leaves within the canopy (data not shown). • Litter N concentration was always greater in fertilized plots (b), but more so when the last of the fertilized leaves began to fall, potentially due to differences in timing of green leaf production or senescence. Total litter N content was greater in the fertilized plots (not shown). Introduction: Globally, the atmospheric concentration of CO2 is rising, and this has the potential to increase forest productivity and carbon storage. However, forest response to atmospheric CO2 enrichment has long been thought to be modified by nitrogen (N) availability. CO2-mediated increases in forest productivity, and corresponding increases in N demand, have been predicted to exacerbate any existing N limitation within an ecosystem (Figure 1). In particular, the immobilization of N in plant biomass or soil carbon may provide a negative feedback to plant growth by further decreasing soil N availability. • Forest response to elevated [CO2] depends on the potential for an ecosystem to meet productivity and carbon storage demands. If soil N is limiting, PNL may be inevitable, though plants can compensate for N deficiency for a short time through changes in plant and soil C:N ratios and NUE, and N mining via increased fine root growth. • In order to predict the role of rising [CO2] on future carbon storage in forested ecosystems, it is necessary to first determine whether a forested ecosystem is limited by the availability of N. This requires the addition of excess N, where predicted responses may include: • Increased leaf [N] • Increased stem production • Increased LAI • Decreased root proliferation (b) Figure 1: Adapted from Luo et al. 2004, where they have termed the process by which elevated [CO2] causes the available N in an ecosystem to be allocated to long-lived plant biomass pools (pathway 1) or immobilized to increased soil carbon stocks (pathway 2), progressive N limitation (PNL). PNL depends both on a CO2-mediated stimulation of net primary productivity and the related increase in N demand, both of which have been observed in the CO2-enriched plots at ORNL FACE. Methods: The on-going ORNL (Oak Ridge National Lab) FACE (Free-Air CO2 Enrichment) experiment was initiated in 1998 in a 15-m tall, 10-year old, closed-canopy sweetgum stand. The FACE experimental design consists of five 25-m diameter plots. FACE apparatus were installed in four of the plots (Figure 2a), and two FACE rings blow elevated concentrations of atmospheric CO2 (approximately 550 ppm). A fertilization experiment was initiated in a sweetgum stand adjacent to ORNL FACE (Figure 2a,b) in 2004, when the trees were approximately 19-m tall. The trees were initially planted at the same time and in the same manner as those in the FACE experiment, and the stand encompasses historical FACE rings 6 and 7. The plantation was fertilized with 200 kg ha-1 of N as urea in March of 2004 and again in March of 2005 before leaf-out, in a replicated, randomized complete block design (Figure 2b). I measured several productivity parameters within the fertilization experiment, including tree annual basal area increment (BAI, cm2 m-2), fine root productivity, and leaf litterfall. Leaf [N] and canopy N distribution were also measured by sampling from several heights within the canopy, and felling trees from within each experimental plot. Resin access tubes were used to assess fertilization effects on N availability (Figure 3). Summary: Increases in stem and leaf production, and leaf N concentration, in response to N addition support the conclusion that stem production in this sweetgum plantation is limited by the availability of N. It is likely that stem production in ORNL FACE is also limited by N availability, which has implications for future forest carbon storage in response to rising atmospheric [CO2] (seeFuture Research). Canopy response to N addition was dependent on canopy height and the timing of leaf production, and stem production response was dependent on initial stem basal area. Fine-root responses to N addition are currently being examined. References: Luo, Y., B. Su, W.S. Currie, J.S. Dukes, A. Finzi, U. Hartwig, B. Hungate, R.E. McMurtrie, R. Oren, W.J. Parton, D.E. Pataki, M.R. Shaw, D.R. Zak, and C.B. Field. 2004. Progressive nitrogen limitation of ecosystem responses to rising atmospheric carbon dioxide. Bioscience54: 731-739. Norby, R.J., J. Ledford, C.D. Reilly, N.E. Miller, and E.G. O’Neill. 2004. Fine-root production dominates response of a deciduous forest to atmospheric CO2 enrichment. PNAS101: 9689-9693. Scott, D.A., J.A. Burger, D.J. Kaczmarek, and M.B. Kane. 2004. Growth and nutrition response of young sweetgum plantations to repeated nitrogen fertilization on two site types. Biomass & Bioenergy27: 313-325. • Future research: • According to Luo et al. (2004), limited soil N availability will eventually limit forest productivity response to elevated [CO2]. One of the main premises of PNL theory is that increases in whole plant N are due to N sequestration in plant biomass (pathway 1). However, previous research at ORNL FACE (Norby et al. 2004) has shown that the extra carbon fixed and N taken up is not being stored in long-lived plant biomass, such as wood, but instead much of the N in the CO2-enriched plots is invested in fine root productivity, a pool which has a fast turnover time. • Future research involving 15N-enrichment and fine-root decomposition techniques will concentrate on • the unanswered questions regarding interactions between carbon and N cycling belowground in ORNL FACE (pathway 2): • Will CO2-mediated increases in fine-root litter stimulate microbial activity and increase decomposition rates? • Are decomposition rates dependent on the fine-root length and N content within different root diameter classes? • Will CO2-mediated increases in labile carbon substrate for microbial growth increase microbial N demand and immobilization? *Research supported by US Department of Energy Office of Science, Biological and Environmental Research.






















