Download
1 / 75
750 likes | 805 Views
Learn about the three types of muscle tissue, their structures, organization, and development. Explore the terminology and cellular characteristics of skeletal, cardiac, and smooth muscles.

E N D
Muscle Tissue DEC 2015
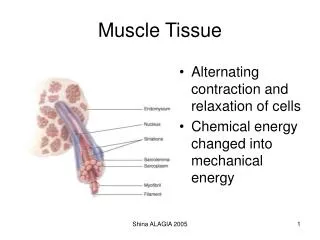
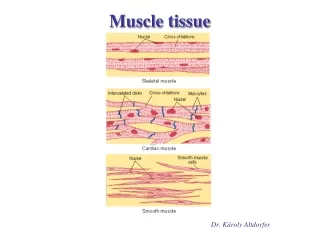
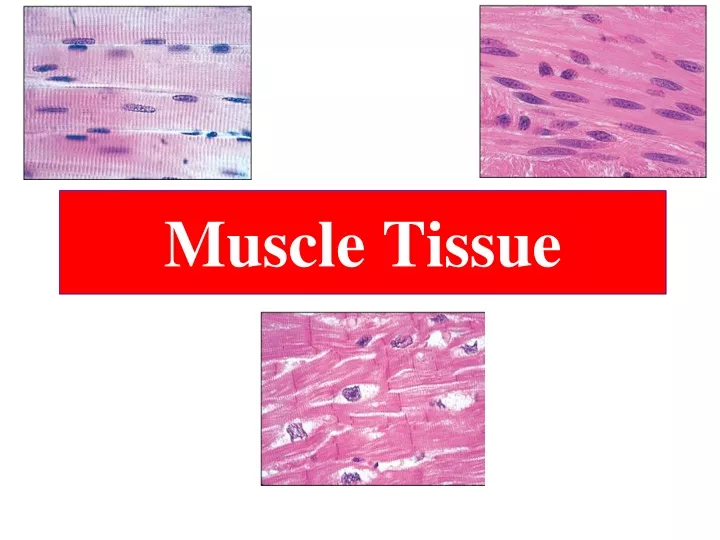
Muscle tissue • TERMINOLOGY • Sarco & Myo • Myofiber • Muscle fiber • Sarcolemma • Sarcoplasm • Sarcoplasmic Reticulum • Differentiated cells containing contractile proteins • Most muscle cells: Mesodermal origin • Three types of cells on the basis of morphological and functional characteristics: 1. Skeletal M. -Very long , cylindrical multinucleated cells, with Cross striations - Voluntary, quick, forceful contraction 2. Cardiac M. - Elongated, branched cells, with Cross striations - Involuntary , vigorous and rhythmic contraction 3. Smooth M. - Fusiform cells, No Cross striation - Slow, Involuntary contraction
Figure 10—1. Structure of the 3 muscle types. The drawings at right show these muscles in cross section. Skeletal muscle is composed of large, elongated, multinucleated fibers. Cardiac muscle is composed of irregular branched cells bound together longitudinally by intercalated disks. Smooth muscle is an agglomerate of fusiform cells. The density of the packing between the cells depends on the amount of extracellular connective tissue present.
The three types of muscle. Light micrographs of each type, accompanied by labeled drawings. (a):Skeletal muscle is composed of large, elongated, multinucleated fibers that show strong, quick, voluntary contractions. (b):Cardiac muscle is composed of irregular branched cells bound together longitudinally by intercalated disks and shows strong, involuntary contractions. (c): Smooth muscle is composed of grouped, fusiform cells with weak, involuntary contractions. The density of intercellular packing seen reflects the small amount of extracellular connective tissue present.
Development of skeletal muscle. Skeletal muscle begins to differentiate when mesenchymal cells called myoblasts align and fuse together to make longer, multinucleated tubes called myotubes. Myotubes synthesize the proteins to make up myofilaments and gradually begin to show cross striations by light microscopy. Myotubes continue differentiating to form functional myofilaments and the nuclei are displaced against the sarcolemma. Part of the myoblast population does not fuse and differentiate, but remains as a group of mesenchymal cells called muscle satellite cells located on the external surface of muscle fibers inside the developing external lamina. Satellite cells proliferate and produce new muscle fibers following muscle injury.
Organization of skeletal tissue • Epimysium, Perimysium, Endomysium • One of the roles of connective tissue: - mechanically transmit the forces generated by contracting muscle cells • A rich capillary network
Figure 10. Structure and function of skeletal muscle. The drawing at right shows the area of muscle detailed in the enlarged segment. Color highlights endomysium, perimysium, and epimysium.
Figure 10—3. Cross section of striated muscle stained to show collagens type I and III and cell nuclei. The endomysium is indicated by arrowheads and the perimysium by arrows. At left is a piece of epimysium. Picrosirius-hematoxylin stain. High magnification.
Figure 10—4. Cross section of striated muscle immunohistochemically stained for laminin, a protein component of the endomysium, which appears in various shades of brown. In the upper right corner is a slightly oblique section of a small nerve. Laminin is also present around nerve fibers.
Figure.5. Longitudinal section of striated muscle fibers. The blood vessels were injected with a plastic material before the animal was killed. Note the extremely rich network of blood capillaries around the muscle fibers. Giemsa stain. Photomicrograph of low magnification made under polarized light.
Fig.6. Striated skeletal muscle in longitudinal section (lower) and in cross section (upper). The nuclei can be seen in the periphery of the cell, just under the cell membrane, particularly in the cross sections of these striated fibers. H&E stain. Medium magnification.
Fig.7. Striated skeletal muscle in longitudinal section. In the left side of the photomicrograph the insertion of collagen fibers with the muscle is clearly seen. Picrosirius—polarized light (PSP) stain. Medium magnification.
Organization of skeletal muscle fibers • Cross-striation of alternating light and dark bands • A bands (Anisotropic) • I bands (Isotropic) • Z line • Sacomere : 2.5 µm in resting muscle • A rich capillary network
-Myofibrils: The sarcoplasm is filled with long cylindrical filamentous bundles called myofibrils (1-2 µm) • -Sarcomere pattern due to the presence of two types of filaments: • 1. Thick & 2. Thin ( both 55% protein) • - Thick : • - 1.6 µm long , 15nm wide • - Occupy the A band (central portion) • -Thin : • - 1.0µm long , 8 nm wide • - BTN thick filaments • - One end attached to the Z line • - I bands consist of the portions of the thin filaments • Fig.11. Structure and position of the thick and thin filaments in the sarcomere. The molecular structure of these components is shown at right.
Fig.8. Longitudinal section of skeletal muscle fibers. Note the dark-stained A bands and the light-stained I bands, which are crossed by Z lines. Giemsa stain. High magnification.
Fig.9. Skeletal muscle in longitudinal section. Note the striation in the muscle cells and the moderate amount of collagen (yellow). PSP stain. High magnification.
-A band (central portion): • - mainly of thick filaments + portions of overlapping thin filaments • -I bands • - consist of the portions of the thin filaments • H bands • - The lighter zone of the center of A band • - Consisting only of the rod-like portions of the myosin molecule • - M line • - Bisecting the H band • - Lateral connections BTN thick filaments, • -Consisting of 2 main proteins: • 1-Myomesin: a myosin-binding protein, holds myosin in place • 2-Creatine kinase is the major protein • - Transfer of a phosphate group from phosphocreatine to ADP • - Hexagon : each thick surrounded by 6 thin
Myosin-binding site • Fig.13. Schematic representation of the thin filament, showing the spatial configuration of 3 major protein components–actin, tropomyosin, and troponin. The individual components in the upper part of the drawing are shown in polymerized form in the lower part. The globular actin molecules are polarized and polymerize in one direction. Note that each tropomyosin molecule extends over 7 actin molecules. TnI, TnC, and TnT are troponin subunits.
Filaments structure • Myosin: consist of: • -Two identical heavy chains: • Thin, Rod-like, myosin tail • -Two pairs light chains : • Small globular projections, heads • -Enzymatic capacity to hydrolyze ATP • (ATPase activity) & actin binding ability • α-actinin, a major component of the Z line, to anchor actin filaments to the region • α-actinin, and Desmin , to tie adjacent sarcomeres together, keeping the microfibrils in register • Tropomyosin • Troponin, TnT, TnC, TnT
Fig.12. Transverse section of skeletal muscle myofibrils illustrating some of the features diagrammed in Fig.11. • I, I band; • A, A band; • H, H band; • Z, Z line. x36,000.
Filaments structure • Nebulin, attached to Z line , parallel of actin, and regulate the actin length during the evolution of muscle (Involve in myogenesis) • Myomesin, lateral attachment of myosin in M line, a myosin-binding protein, holds myosin in place • C- protein , Cross attachment of myosin in M line • Titin, attachment of Myosins to Z line, maintaining the central position
Sarcoplasmic Reticulum & Transverse tubule system • Transverse (T) tubule • Terminal Cisternae • Triad
Fig.10. Electron micrograph of skeletal muscle of a tadpole. Note the sarcomere with its A, I, and H bands and Z line. The position of the thick and thin filaments in the sarcomere is shown schematically in the lower part of the figure. As illustrated here, triads in amphibian muscle are aligned with the Z line in each sarcomere. In mammalian muscle, however, each sarcomere exhibits 2 triads, one at each A—I band interface (see Fig.16). x35,000.
Figure 10—15. Electron micrograph of a transverse section of fish muscle, showing the surface of 2 cells limiting an intercellular space. Note the invaginations of the sarcolemma, forming the tubules of the T system (arrows). The dark, coarse granules in the cytoplasm (lower left) are glycogen particles. The section passes through the A band (upper right), showing thick and thin filaments. The I band is sectioned (lower left), showing only thin filaments. x60,000.
Fig.16. Electron micrograph of a longitudinal section of the skeletal muscle of a monkey. Note the mitochondria (M) between adjacent myofibrils. The arrowheads indicate triads–2 for each sarcomere in this muscle–located at the A—I band junction. A, A band; I, I band; Z, Z line. x40,000.
Fig.17. Segment of mammalian skeletal muscle. The sarcolemma and muscle fibrils are partially cut, showing the following components: The invaginations of the T system occur at the level of transition between the A and I bands twice in every sarcomere. They associate with terminal cisternae of the sarcoplasmic reticulum (SR), forming triads. Abundant mitochondria lie between the myofibrils. The cut surface of the myofibrils shows the thin and thick filaments. Surrounding the sarcolemma are a basal lamina and reticular fibers.
Mechanism of contraction Fig.14. Muscle contraction, initiated by the : 1. Binding of Ca2+ to the TnC unit of troponin, which exposes the myosin binding site on actin (cross-hatched area). 2. In a second step, the myosin head binds to actin and the ATP breaks down into ADP, yielding energy, which produces a movement of the myosin head. 3. As a consequence of this change in myosin, the bound thin filaments slide over the thick filaments. This process, which repeats itself many times during a single contraction, leads to a complete overlapping of the actin and myosin and a resultant shortening of the whole muscle fiber. I, T, C are troponin subunits.
Innervation • Myelinated motor nerves • within the perimysial connective tissue branch out • At the site of innervation: 1. The nerve loses its myelin sheath 2. A dilated termination forms 3. Sits within a trough on the muscle cell surface 4. The motor end plate(MEP) or myoneural junction forms - Axon terminal is covered by a thin cytoplasmic layer of Schwann cell - Mitochondria & Synaptic vesicles containing Acetyl choline - Synaptic cleft -Junctional folds
Fig.18. Ultrastructure of the motor end-plate and the mechanism of muscle contraction. The drawing at the upper right shows branching of a small nerve with a motor end-plate for each muscle fiber. The structure of one of the bulbs of an end-plate is highly enlarged in the center drawing. Note that the axon terminal bud contains synaptic vesicles. The region of the muscle cell membrane covered by the terminal bud has clefts and ridges called junctional folds. The axon loses its myelin sheath and dilates, establishing close, irregular contact with the muscle fiber. Muscle contraction begins with the release of acetylcholine from the synaptic vesicles of the end-plate. This neurotransmitter causes a local increase in the permeability of the sarcolemma. The process is propagated to the rest of the sarcolemma, including its invaginations (all of which constitute the T system), and is transferred to the sarcoplasmic reticulum (SR). The increase of permeability in this organelle liberates calcium ions (drawing at upper left) that trigger the sliding filament mechanism of muscle contraction. Thin filaments slide between the thick filaments and reduce the distance between the Z lines, thereby reducing the size of all bands except the A band. H, H band; S, sarcomere.
Figure 10–13.The neuromuscular junction (NMJ). Before it terminates in a skeletal muscle, each motor axon bundled in the nerve forms many branches, each of which forms a synapse with a muscle fiber. (a): Silver staining can reveal the nerve bundle (NB), the terminal axonal twigs, and the motor end plates (MEP) on striated muscle fibers (S). X1200. (b): A SEM shows the branching ends of a motor axon, each covered by an extension of the last Schwann cell and expanded terminally as a motor end plate embedded in a groove in the external lamina of the muscle fiber. (c): Diagram indicating key features of a typical neuromuscular junction: synaptic vesicles of acetylcholine (ACh), a synaptic cleft, and a postsynaptic membrane. This membrane, the sarcolemma, is highly folded to increase the number of Ach receptors at the NMJ. Receptor binding initiates muscle fiber depolarization, which is carried to the deeper myofibrils by the T tubules.
Innervation • Motor unit • Muscle spindles - Intrafusal fibers (Modified muscle fibers) - The sensory nerve fibers penetrate the muscle spindles -Nuclear bag Nuclear chain • Golgi tendon organ -In Tendons, near the insertion sites of muscle fibers, a connective tissue sheath encapsulates several large bundles of collagen fibers that are continuous with the collagen fibers that make up the myotendinous junction. - Sensory nerves penetrate the capsule---------forms Golgi Tendon Organs
Fig.19. Muscle spindle showing afferent and efferent nerve fibers that make synapses with the intrafusal fibers (modified muscle fibers). Note the complex nerve terminal on the intrafusal fibers. • The two types of intrafusal fibers, one with a small diameter and the other with a dilation filled with nuclei, are shown. Muscle spindles participate in the nervous control of body posture and the coordinate action of opposing muscles.
Figure 10–14.Sensory receptors associated with skeletal muscle.(a): Diagram shows both a muscle spindle and a tendon organ. Muscle spindles have afferent sensory and efferent motor nerve fibers associated with the intrafusal fibers, which are modified muscle fibers. The size of the spindle is exaggerated relative to the extrafusal fibers to show better the nuclei in the intrafusal fibers. (b): TEM cross—section near the end of a muscle spindle shows the capsule (C), sensory myelinated axons (MA), and the intrafusal muscle fibers (MF). These thin fibers differ from the ordinary skeletal muscle fibers in having essentially no myofibrils. Their many nuclei can either be closely aligned (nuclear chain fibers) or piled in a central dilatation (nuclear bag fibers). Satellite cells (SC) are also present within the external lamina of intrafusal fibers. Muscle spindles detect contraction of neighboring (extrafusal) muscle fibers during body movement and participate in the nervous control of body posture and the coordinate action of opposing muscles. The tendon organ collects information about the degree of tension among tendons and relays this data to the CNS, where the information is processed with that from muscle spindles to protect myotendinous junctions and help coordinate fine muscular contractions.
Fig.20. Drawing of a Golgi tendon organ. This structure collects information about differences in tension among tendons and relays data to the central nervous system, where they are processed and help to coordinate fine muscular contractions.
System of Energy production • To forms of stored available energy in skeletal muscle: 1. ATP 2. Phosphcreatine 3. Chemical energy in glycogen depots • Classification of muscle fibers( morphological, Histochemical, Biochemical) 1. Type I (Slow fibers): Red fibers: High myoglobin: Continuous contraction, energy from oxidative phosphorylation of fatty acids - Long time activities: Muscles of migrant birds, deep-diving ocean mammals (whales, seals), 2. Type II (Fast fibers): white fibers: Less Myoglobin: Rapid and discontinuous contraction, glycolysis as source of energy, - Chicken muscles, External muscles of eye 3. Intermediate: Mixed characteristics: Most human muscles
Fig.21. Section of tongue, an organ rich in striated skeletal muscle fibers. These fibers appear brown because the section was immunohistologically stainedto show myoglobin. The light-colored areas among and above the muscle fibers contain connective tissue. In the upper region of the section, stratified and cornified epithelium can be seen. Nuclei are stained by hematoxylin.
Cardiac Muscle - Cells are within a chain, bifurcate, or branch, and bind to cells in adjacent chains - A mature cardiac muscle cell: - 15 µm diameter - 85-100 µm length -Cross striation pattern - Only one or two Centrally located pale-staining Nuclei - Endomysium containing a rich capillary network -Unique characteristic: Intercalated disks - Junctional complexes - Two regions 1. Transverse portion : Fascia adherentes, Macula adherentes (Desmosomes) 2. Longitudinal portion: Gap junctions
Figure 10—22. Drawing of a section of heart muscle, showing central nuclei, cross-striation, and intercalated disks.
Fig.23. Photomicrograph of cardiac muscle. Note the cross-striation and the intercalated disks (arrowheads). Pararosaniline—toluidine blue (PT) stain. High magnification.
Cardiac Muscle -Intercalated disks Two regions 1. Transverse portion - Runs across the fibers a. Fascia adherentes - Serve as anchoring sites for actin filaments -(Hemi-Z bands) b. Macula adherentes (Desmosomes) - Bind the cardiac cells together -Preventing them from pulling apart under constant contractile activity 2. Longitudinal portion -Gap junctions
Fig.24. Longitudinal section of portions of 2 cardiac muscle cells. The transversely oriented parts of the intercalated disk consist of a fascia adherens and numerous desmosomes. The longitudinal parts (arrows) contain gap junctions. Mitochondria (M) are numerous. Fibrils of reticular fibers are seen between the two cells. x18,000.