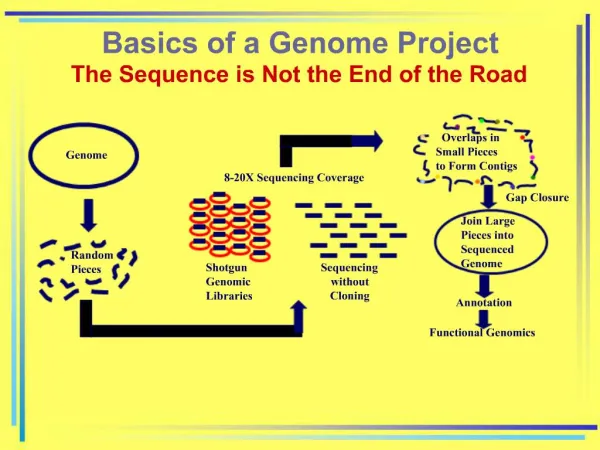
Download

1 / 53
570 likes | 797 Views
Genome Rearrangement. By Ghada Badr Part II. Genome Models. Genomes can be modeled by each gene can be assigned a unique number and is exactly found once in the genome. permutations :.

E N D
Genome Rearrangement By Ghada Badr Part II
Genome Models • Genomes can be modeled by each gene can be assigned a unique number and is exactly found once in the genome. permutations: • Signed Permutation: Each gene may be assigned + or - sign to indicate the strand it resides on. • Unsigned Permutation: If the corresponding strand is unknown.
Genome Rearrangement Our problem: Given a set of genomes and a set of possible evolutionary events (operations), find a shortest set of events transforming those genomes into one another. What are the Rearrangement events (Operation)?
Rearrangement Operations Rearrangement operations affect gene order and gene content. There are various types: In case of single-chromosome genome: • Inversions • Transpositions • Reverse transpositions • Gene Duplications • Gene loss In case of multiple-chromosomes genomes we add: • Translocations • fusions • fissions
Rearrangement Problems Our problem: Given a set of genomes and a set of possible evolutionary events (operations), find a shortest set of events transforming those genomes into one another. Any set of operations yields a distance between genomes, by counting the minimum number of operations needed to transform one genome into the other.
Rearrangement Problems Our problem: Given a set of genomes and a set of possible evolutionary events (operations), find a shortest set of events transforming those genomes into one another. Two classical problems Computing the distance d() Computing one optimal sorting sequence of events.
Rearrangement Operations Can we have a unifying framework in which circular and linear chromosomes can coexist throughout evolving genomes? Can we have a unifying view of Genome Rearrangements? (Bergeron 2006) A Double Cut and Join Operation DCJ was introduced.
Rearrangement Operations - DCJ • Double Cut-and-Join DCJ was first proposed by Yancopoulos et. al. (2005). • Allows to model all the classical operations (inversions, translocations, fissions, fusions, transposition, and block interchanges) with a single operation. • This general model accounts for the genomic evidence of the coexistence of both linear and circular chromosomes in many genomes. • Both the DCJ sorting and distance problems can be solved in O(n) time by Bergeron et. al. (2006)
Rearrangement Operations - DCJ • A genea is an oriented sequence of DNA that starts with a tailatand ends with a headah. • Two consecutive genes do not necessarily have the same orientation, thus adjacency of two consecutive genes a and b, can be of four different types: {ah,bt},{ah,bh},{at,bt},{at,bh} , , , • An extremity that is not adjacent to any other gene is called telomeres by a singleton set {ah} or {at}. • We can use adjacencies to represent both genomes with multiple or uni-chromosomes.
Rearrangement Operations - DCJ • A genome is a set of adjacencies and telomeres such that the tail or head of any gene appears in exactly one adjacency or telomere. Replace each gene by two extremities Example at ah ct ch dh dt bt bh et eh ft fh gt gh Genome A: chr1: a c -d chr2: b e chr3: f g Adjacencies: {ah, ct}{ch, dh} {bh, et} {fh, gt} Telomere:{at} {dt} {bt} {eh}{ft}{gh} A = {{at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh}}
Rearrangement Operations - DCJ • DCJ operations: {p,q}{r,s} {p,r}{s,q} or { p,s} {q,r} a)
Rearrangement Operations - DCJ • DCJ operations: {p,q}{r} {p,r}{q} or{p}{q,r} b)
Rearrangement Operations - DCJ • DCJ operations: {q} {r} {q,r} c)
Rearrangement Operations - DCJ • DCJ operations: Example: Genome A: chr1: a c -d chr2: b e chr3: f g Adjacencies and telomeres are: {ah, ct}{ch, dh} {bh, et} {fh, gt} {at} {dt} {bt} {eh}{ft}{gh} {ah,ct}{fh, gt} -->{ah,fh}{ct,gt} Genome A: chr1: a -f chr2: b e chr3: d -c g {ah,ct}{fh, gt} -->{ah,gt}{ct,fh} Genome A: chr1: a g chr2: b e chr3: f c -d
DCJ sorting and Distance problems Problem: Given two genomes A and B defined on the same set of genes, find a shortest sequence of DCJ operations that transforms A into B. The length of such a sequence is called the DCJ distance between A and B, dcj(A,B).
DCJ sorting and Distance problems Example: Replace each gene by two extremities at ah ct ch dh dt bt bh et eh ft fh gt gh Genome A: chr1: a c -d chr2: b e chr3: f g at ah bt bh ct ch dt dh et eh ft fh gt gh Genome B: chr 1: a b c d chr 2: e f g Get adjacencies and telomeres for each genome: A= {{ah, ct}{ch, dh} {bh, et} {fh, gt} {at} {dt} {bt} {eh}{ft}{gh}} B = {{at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh}}
DCJ sorting and Distance problems Greedy Algorithm to sort by DCJ: Genome A: chr1: a c -d chr2: b e chr3: f g {ah, ct}{ch, dh} {bh, et} {fh, gt} {at} {dt} {bt} {eh}{ft}{gh} {ah, bt}{ch, dh} {bh, et} {fh, gt} {at} {dt} {ct} {eh}{ft}{gh} Genome A: chr1: a b e chr2: c -d chr3: f g {ah, bt} {ch, dh} {bh, ct} {fh, gt} {at} {dt} {et} {eh}{ft}{gh} Genome A: chr1: a b c -d chr2: e chr3: f g {ah, bt} {ch, dt} {bh, ct} {fh, gt} {at} {dh} {et} {eh} {ft}{gh} Genome A: chr1: a b c d chr2: e chr3: f g {at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh} Genome B: chr1: a b c d chr2: e f g
DCJ sorting and Distance problems Optimal and O(n) time.
DCJ sorting and Distance problems Adjacency Graph (bipartite graph): {ah, ct}{ch, dh} {bh, et} {fh, gt} {at} {dt} {bt} {eh}{ft}{gh} {at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh} Vertices: adjacencies and telomeres Edges: connect an edge from A to B between adjacencies or telomers that have common elements. Graph can be easily constructed in O(n) time and space
DCJ sorting and Distance problems Adjacency Graph (bipartite graph): IF SORTED {at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh} {at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh} In each iteration: the algorithm increments C by one or I by two When sorted: n = C + I/2 dcj(A,B) n
1 cycle 4odd paths 1 even path DCJ sorting and Distance problems Adjacency Graph (bipartite graph): {ah, ct}{ch, dh} {bh, et} {fh, gt} {at} {dt} {bt} {eh}{ft}{gh} {at}{ah, bt}{bh, ct}{ch, dt}{dh} {et} {eh,ft} {fh,gt} {gh} dcj(A,B) = n - (cycles + oddPath/2) = 7-1-4/2 = 4
Genome Rearrangement and phylogeny • Genome rearrangements events are rare, these changes of gene orders enable biologists to reconstruct histories far back in time. • Extend the notion of genome rearrangement distance to the optimal positioning of Steiner points in the appropriate space of a given distance metric. • Two phylogenetic versions of the Steiner Problem (the first inside the other): • Inner problem: optimizing internal nodes of a given tree, where n leaves are labeled. • Outer problem: optimizing over all trees with n leaves.
Genome Rearrangement and phylogeny • We will discuss the inner problem defined as follows: Given a fixed phylogeny (tree) T, together with a set of K permutations (genome), each of size n corresponding to the terminal (leaf) nodes. Find a set of permutations corresponding to the internal nodes such that the total weight w(T) is minimized, where w(T) is defined as: w(T) = ∑ d(x,y) for all (x,y) in T Here d(.,.) is the genome rearrangement distance metric defined on pairs of permutations.
Genome Rearrangement and phylogeny • Consider a heuristic for the problem of computing the internal nodes, where T is a star on three vertices. • We will study a more basic problem, the median problem. • Divide the problem on an arbitrary binary tree into a number of overlapping median problems and apply the median algorithm iteratively to search for a heuristic solution to the original problem. • internal nodes retain biological meaning, and edges represent transitions between states of genome.
Median Problem • The median-based method for phylogeny reconstruction was first proposed by Sankoff and Blanchette (1998). • The idea is to build the global solution by aggregating local solutions for the simplest problem: Find a Steiner point M of three genomes. • After an initialization step, the algorithm iterates over a tree, repeatedly resetting the permutations of internal nodes to the medians of their three neighbors. Continue till a convergence occurs.
Median Problem • The median of three or simply the median problem: Find a permutation such that the sum of distances is minimized between and each of the starting permutation = {}. • Find a permutation M that minimizes the median score S(), where: S() = d1, M + d2,M + d3,M
Median Problem Constructing phylogeny from medians
Median Problem • The median problem: Find a permutation such that the sum of distances is minimized between and each of the starting permutation = {}. • What are the distance measures that we can use? • Distances: breakpoint, reversal … • A breakpoint median has no straightforward biological interpretation and they are not unique. • Breakpoint medians score poorly compared to reversal medians.
Reversal Median • Reversal median Problem: Find a solution to the median problem using the reversal distance. • Find a permutation such that the sum of reversal distances is minimized between and each of the starting genomes. • The reversal median is NP-hard problem. • Why?
Reversal Median Reversal graph for n = 3 Vertices: all permutations of n = 3. Edges: connect an edge between 1 and 2 if reversal distance d(1, 2) = 1.
Reversal Median Reversal graph for n = 3 • distance d(i, k) = shortest path between v1 and v2. • Finding the median is equivalent to finding the minimum Steiner tree for the graph.
Reversal Median Reversal graph for n = 3 • The graph is huge |V| = n!.2n • A feasible graph-search algorithm is not possible! • What technique we can use to develop an algorithm for this kind of problems?
Reversal Median • We will study a branch-and-bound algorithm by Adam Siepel 2001. • This algorithm depends only on the availability of a rapidly computable distance metric.
Reversal Median • The median score S() of a set of equally sized permutations = {}, separated by distances d1,2, d1,3, and d2,3, obeys these bounds: d1,2 + d1,3+ d2,3 S() min { (d1,2+d2,3),(d1,2+d1,3), (d2,3+d1,3)} 2
Reversal Median • Assume that is in the shortest path between and the median M, and is separated from by distances d1,, d1,, and d2,, the median score S() d2, + d3,+ d2,3 d1,+ S()d1,+min{(d2,+d3,),(d3,+d2,3), (d2,3+d2,)} 2
Reversal Median Algorithm (sketch): • Establish upper and lower bounds using a rapid reversal distance algorithm, Mmin and Mmax. • Start with one of the three permutations, say . • Assume the median is M = . • Push the corresponding vertex v in a priority stack s for the best scoring vertices. • While s is not empty • Pop the most promising vertex v from s. • If best score of v Mmax then stop • Generate all possible vertices that can be obtained from v by single reversal. • For each possible unmarked • Calculate bound for the previous equation min, max. • If max= Mmin then M = and stop. (median is found) • Add to stack s only if max< Mmax (pruning) • update Mmax= max if max< Mmax . • End for loop. • End while loop.
Reversal Median Algorithm (sketch): • Establish upper and lower bounds using a rapid reversal distance algorithm, Mmin and Mmax. • Start with one of the three permutations, say . • Assume the median is M = . • Push the corresponding vertex v in a priority stack s for the best scoring vertices. • While s is not empty • Pop the most promising vertex v from s. • If best score of v Mmax then stop • Generate all possible vertices that can be obtained from v by single reversal. • For each possible unmarked • Calculate bound for the previous equation min, max. • If max= Mmin then M = and stop. (median is found) • Add to stack s only if max< Mmax (pruning) • update Mmax= max if max< Mmax . • End for loop. • End while loop.
Reversal Median Algorithm (sketch): • Establish upper and lower bounds using a rapid reversal distance algorithm, Mmin and Mmax. • Start with one of the three permutations, say . • Assume the median is M = . • Push the corresponding vertex v in a priority stack s for the best scoring vertices. • While s is not empty • Pop the most promising vertex v from s. • If best score of v Mmax then stop • Generate all possible vertices that can be obtained from v by single reversal. • For each possible unmarked • Calculate bound for the previous equation min, max. • If max= Mmin then M = and stop. (median is found) • Add to stack s only if max< Mmax (pruning) • update Mmax= max if max< Mmax . • End for loop. • End while loop.
Reversal Median Algorithm (sketch): • Establish upper and lower bounds using a rapid reversal distance algorithm, Mmin and Mmax. • Start with one of the three permutations, say . • Assume the median is M = . • Push the corresponding vertex v in a priority stack s for the best scoring vertices. • While s is not empty • Pop the most promising vertex v from s. • If best score of v Mmax then stop • Generate all possible vertices that can be obtained from v by single reversal. • For each possible unmarked • Calculate bound for the previous equation min, max. • If max= Mmin then M = and stop. (median is found) • Add to stack s only if max< Mmax (pruning) • update Mmax= max if max< Mmax . • End for loop. • End while loop.
Reversal Median Algorithm (sketch): • Establish upper and lower bounds using a rapid reversal distance algorithm, Mmin and Mmax. • Start with one of the three permutations, say . • Assume the median is M = . • Push the corresponding vertex v in a priority stack s for the best scoring vertices. • While s is not empty • Pop the most promising vertex v from s. • If best score of v Mmax then stop • Generate all possible vertices that can be obtained from v by single reversal. • For each possible unmarked • Calculate bound for the previous equation min, max. • If max= Mmin then M = and stop. (median is found) • Add to stack s only if max< Mmax (pruning) • update Mmax= max if max< Mmax . • End for loop. • End while loop.
Reversal Median Algorithm (sketch): • Establish upper and lower bounds using a rapid reversal distance algorithm, Mmin and Mmax. • Start with one of the three permutations, say . • Assume the median is M = . • Push the corresponding vertex v in a priority stack s for the best scoring vertices. • While s is not empty • Pop the most promising vertex v from s. • If best score of v Mmax then stop • Generate all possible vertices that can be obtained from v by single reversal. • For each possible unmarked • Calculate bound for the previous equation min, max. • If max= Mmin then M = and stop. (median is found) • Add to stack s only if max< Mmax (pruning) • update Mmax= max if max< Mmax . • End for loop. • End while loop. O(n3d) with d = min{d1,2 + d1,3+ d2,3} With faster average running time
Conclusions • Described Double Cut and Join DCJ operation: A unifying view of genome rearrangements. • Presented a branch and bound median-based approach for building phylogeny using reversal distance. • Many other problems in genome rearrangement as “Genomehalving problem”
Genome Halving a f c d f b a f c b g g a e d e g e d b c
Genome Halving Duplication a f c d f b a f c b g g a e d e g e d b c
b b c c d d e e g g a a f f Genome Halving a f c d f b a f c b g g a e d e g e d b c
b b c c d d e e g g a a f f Genome Halving a f c d f b a f c b g g a e d e g e d b c
b b b c c c d d d e e e g g g a a a f f f Genome Halving a f c d f b a f c b g g a e d e g e d b c
b c d e g a f Genome Halving a f c b g e d
b b b c c c d d d e e e g g g a a a f f f Genome Halving a f c b g e d
b b b c c c d d d e e e g g g a a a f f f Genome Halving a f c d f b a f c b g g a e d e g e d b c