Download
1 / 9
90 likes | 439 Views
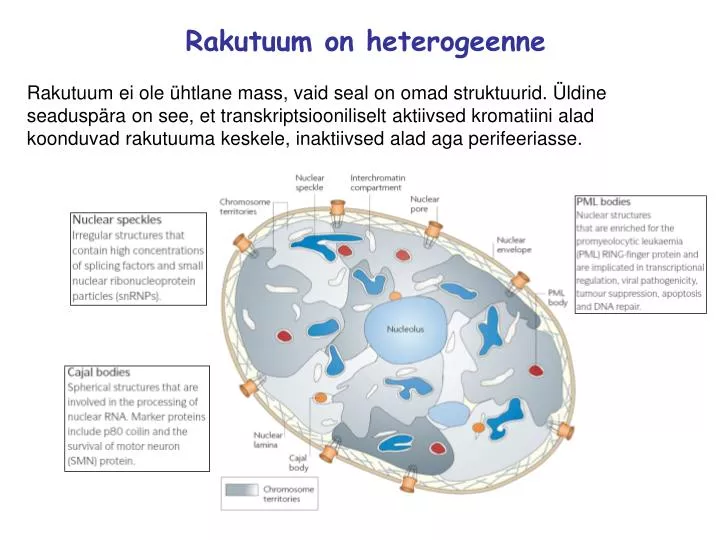
Rakutuum on heterogeenne. Rakutuum ei ole ühtlane mass, vaid seal on omad struktuurid. Üldine seaduspära on see, et transkriptsiooniliselt aktiivsed kromatiini alad koonduvad rakutuuma keskele, inaktiivsed alad aga perifeeriasse. Kromosoomide territooriumid.

E N D
Rakutuum on heterogeenne Rakutuum ei ole ühtlane mass, vaid seal on omad struktuurid. Üldine seaduspära on see, et transkriptsiooniliselt aktiivsed kromatiini alad koonduvad rakutuuma keskele, inaktiivsed alad aga perifeeriasse.
Kromosoomide territooriumid Kromosoomid ei paikne tuumas hajusalt, vaid neil kõigil on oma territoorium. Struktuur eksisteerib ka ühe kromosoomi piires, kuigi selle moodustumise põhimõtted pole päris selged. On teada, et kromosoomi poolt hõivatava ala sees on kanalite võrk, mida mööda pääsevad DNA-le regulaatorvalgud. Geenirikkad kromosoomide alad koonduvad kokku, samuti teevad ka geenivaesed alad ning kromosoomi otsad on alati üksteisest eemal. Kromosoomide omavaheline asetus ruumis võib varieeruda. On küll piirkonnad, kus üks või teine kromosoom eelistatult paikneb, kuid see ei tähenda, et ta oleks igas rakus alati samas kohas. Näiteks T-rakkude diferentseerumise käigus liigub kromosoom 6 rakutuuma keskosast (diferentseerumata rakkudes) tuuma äärealadele (diferentseerunud rakus).
Kromosoomide territooriumid On leitud, et sarnase arengusuunaga rakkudes on kromosoomide ruumiline paiknemine sarnane. Näiteks inimese lümfotsüütides paikneb kromosoom 18 rakutuuma perifeerias ning kromosoom 19 tuuma keskel. Täpselt samasugune on kromosoomide paiknemine ka ahvide lümfotsüütides. Mis on selliste paiknemiste mõte? Arvatavasti aitab aktiivsete geenide koondamine suurematesse “keskustesse” tõsta nende transkribeerimise efektiivsust, samuti aitab represseeritavate alade koondamine hoida need piirkonnad inaktiivsed. Tuntuim “transkriptsioonitehas” on tuumake, kus toimub nii ribosoomide süntees kui ka kõikide rRNA-de transkriptsioon. Ilmselt on olemas sarnased, kuigi väiksemad “tehased” ka RNA polümeraas II geenide transkriptsiooniks. Viimaste aastate uurimused näitavad, et tegelikult toimub genoomis pidevalt enamuse geenide väga väike transkribeerimine. See kehtib ka nende geenide kohta, mille transkriptsioon arvati olevat väga selgelt kas “sees” või “väljas”. Võimalik, et transkriptsiooni “aktivatsioon” tähendab tegelikult geeni liigutamist transkriptsioonikeskusesse ning “repressioon” seda, et geenilookus viiakse sellisest keskusest välja, kuhugi heterokromatiini vahele.
Kromatiini liikumine tuumas Esimesed katsed mõõta kromosoomi ühe lookuse liikuvust rakutuumas andsid tulemuse, et see liigub territooriumil, mille raadius on umbes 0.5-1mm. See on ligikaudu 1% tuuma ruumalast. Nende liikumiste ulatus ja kineetika andsid alust arvata, et tegu on piiratud difusiooniga. Erandiks oli rakutsükli G1 faasi algus, kus sageli nähti lookuse liikumist rohkem kui 2mm ulatuses. Meetodide arenedes jõuti parema lahutuvuseni nii ajas (30 ms) kui ruumis (20 nm) ning siis leiti, et keskmiselt korra minutis teeb fluorestseeruvalt märgistatud lookus umbes 150 nm pikkuse kiire “hüppe”. Sellised hüpped on ATP-st ja temperatuurist sõltuvad, mis näitab, et rakkudes võib eksisteerida aktiivne mehhanism, mis kromatiini liigutab. Ajaliselt on suuremad liikumised piiratud G1 faasi esimese kolmandikuga, peale seda jääb kromatiin suhteliselt paigale, liikudes vaid üsna kitsal territooriumil. Pole teada kuidas kromosoomid oma koha leiavad. On pakutud kahte võimalust: 1. Kromosoomi asukoht tuumas määratakse ära tuuma struktuurvalkude poolt. See variant eeldab, et igat kromosoomi tuntakse spetsiifiliselt ära. 2. Kromosoomid leiavad ise oma koha ning sarnase ekspressioonimustriga piirkonnad koonduvad kokku.
Kromatiini liikumine tuumas On uuritud ka kromatiini liikuvust sõltuvalt selle transkriptsioonilisest aktiivsusest. Selleks tehti rakuliin, kus geenilookusesse olid integreeritud lac repressori seondumisjärjestused. Sellises rakuliinis ekspresseeriti hübriidvalku, milles oli VP16 transkriptsiooni aktivatsioonidomeen ning lac repressori DNA-d siduv domeen. Sõltumatult selle hübriidvalgu olemasolust, liikus märgistatud geenilookus pärast mitoosi rakutuuma keskele. Kuid 3-4 tunni jooksul liikus see tuuma perifeeriasse sel juhul, kui rakkudes ei olnud VP16-lacR hübriidvalku ekspresseeritud. Kui VP16-lacR oli olemas, siis jäi lac repressori seondumisjärjestustega geenilookus rakutuuma keskele. Üldiselt lähevad sellised tulemused kokku paljude teiste andmetega, mis näitavad, et transkriptsiooniliselt aktiivased geenid asuvad tuuma keskosas. Siiski on erandeid, näiteks interferoon-g lookus on alati tuuma perifeerias, sõltumata sellest kas ta on aktiivne või mitte. Samuti on näidatud seda, et hiire b-globiini lookus muutub kõigepealt transkriptsiooniliselt aktiivseks ja alles pärast seda hakkab liikuma tuuma keskossa. Seega ei tähenda tuuma äärealadel paiknemine veel seda, et sellised geenid ei võiks olla aktiivsed. Arvatavasti aitab geenilookuse lokalisatsiooni muutmine eelkõige optimiseerida transkriptsiooni efektiivsust.
Kromatiini liikumine tuumas On ka näiteid, kus väga erineva ekspressioonitasemega geenid paiknevad teineteisele niivõrd lähedal, et nende lokaliseerumine rakutuuma erinevatesse ruumipunktidesse pole võimalik. Näiteks hiire l5-VpreB1 lookus, millest ekspresseeritakse kõikjal ja alati topoisomeraas-3b geeni, asub vaid 1500 bp eemal VpreB1 geenist, mille ekspressioon on väga rangelt pre-B rakkude spetsiifiline. Pärmis on asi üldse veidi erinev. Võrreldes kõrgemate eukarüootidega on pärmis suhteliselt vähe heterokromatiini ning samuti pole seal kromosoomid lokaliseerunud eraldi territooriumitele. Pärmis asuvad tuuma perifeerias nii heterokromatiinsed alad kui ka transkriptsiooniliselt eriti aktiivsed alad. Nimelt on leitud, et tugeva transkriptsiooni korral liigub geenilookus tuumapoori kompleksi lähedale: sealt on hea lühike tee mRNA-d tuumast välja transportida. On selgunud, et mõned tuumapoori moodustavad valgud suudavad ka seonduda aktiivselt transkribeeritavate geenide DNA-ga.
Gene kissing Drosophila geenide repressioon on suuresti sõltuv heterokromatiini moodustamisest PcG (polycomb group) valkude poolt. Heterokromatiini moodustumine algab PcG seondumisega oma spetsiifilsele DNA järjestusele PRE (polycomb response element). Kui ühe PRE-d sisaldava DNA lõigu järjestus kopeeriti veel teisegi kromosoomi, siis need lookused paiknesid ruumiliselt samas kohas. Sarnast erinevate lookuste kolokalisatsiooni on leitud ka transkriptsiooni aktivatsiooni puhul. Näiteks erinevate lõhnade tajumiseks peavad neuronid ekspresseerima ühte umbes 1300-st erinevast lõhnaretseptorist. See saavutakse sellega, et genoomis on vaid üks koopia OR retseptorite (olfactory receptor) transkriptsiooniks vajalik enhancer ning ekspresseerima hakatakse seda OR geeni, millega enhancer ruumiliselt samas punktis kokku saab.
Interaktsioonid kromosoomide vahel Kogu genoomi hõlmavate analüüsidega on püütud selgitada ka seda, et milliste teiste lookustega meid huvitava geeni “X” lookus interakteerub. Selleks kasutatakse 3C meetodi (chromosome conformation capture). Näiteks on sellist katset tehtud hiire Hoxb1 geeniga. Selgus, et enamus Hoxb1 geeni interaktsioone toimus aladega, mis jäid temast kuni 2Mb kaugusele. Samalaadseid katseid on tehtud ka b-globiini lookusega. Ka seal leiti, et enamasti interakteerub b-globiini lookus aladega, mis on temaga samal kromosoomil, kuigi mõnikord kümnete Mb kaugusel. Selgus ka et interaktsioonipartnerid on täiesti erinevad maksarakkudes (kus b-globiini geen on aktiivne) võrreldes ajurakkudega (kus b-globiini geen on inaktiivne). Maksarakkudes oli b-globiini lookus 80% juhtudest koos aktiivselt transkribeeritavate geenidega, samas kui ajurakkudes interakteerus b-globiini lookus 87% juhtumitest selliste aladega, mis polnud aktiivsed.