Download

1 / 102
1.03k likes | 1.38k Views
Chapter 6 mechanisms of genetic recombination. 遗传重组的机制. Main contents. 1. homologous recombination (同源重组) 2. Site-specific recombination (位点专一性重组) 3. Transposition recombination( 转座重组 ) 4. illegitimate recombination( 异常重组 ). 概述:. ◘ 只要有 DNA ,就会发生重组. 减数分裂. 高等生物体细胞核基因、叶绿体和线粒体. 温和噬菌体.

E N D
Main contents • 1. homologous recombination(同源重组) • 2. Site-specific recombination(位点专一性重组) • 3. Transposition recombination(转座重组) • 4. illegitimate recombination(异常重组)
概述: ◘只要有DNA,就会发生重组 减数分裂 高等生物体细胞核基因、叶绿体和线粒体 温和噬菌体 转座子转座 ◘ 生存→变异→突变,重组 损伤修复、适应环境、加速进化
◘重组可分为四类(DNA序列、蛋白质因子) 四种DNA重组的特征
1、homologous recombination(同源重组) (1) Concept homologous or generalized recombination(同源重组或普遍性重组) :联会配对的同源DNA序列之间相互交换对等部分的过程。
(2) 同源重组的特点及影响因素 ①特征: ◘ 涉及同源序列间的联会配对,且交换的片段较大 ◘ 涉及DNA分子在特定的交换位点发生断裂和错接的生化过程异源双链区的生成 ◘ 存在重组热点 ◘ 需要重组酶 ◘ 单链DNA分子或单链DNA末端是交换发生的重要信号
细线期 合线期 ◘ e.g. Euk.减数分裂时的染色单体之间的交换 细菌的转化, 转导,接合,噬菌体重组 粗线期 双线期 终变期
② Influence factors(影响因素) (1)同源区段的长度 同源序列越长越有利于交换 (2) Chromosome structure(染色体结构) (3) Cell types(细胞类型) (4) Sex and age(性别和年龄)
③同源重组的分子机制 A. Chiasmatatypehypothesis(交叉型假说) Janssens(1909年)提出 认为交叉是交换的结果,但并未从分子水平解释重组发生的机制。
Chiasma formation is responsible for generating recombinants.
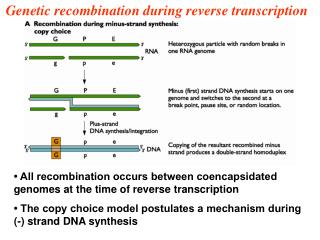
B. Copy choice hypothesis(模板选择学说) • Belling J.首先提出 认为重组(交换)是染色体复制过程中变换模板的结果。
模板选择复制模型存在的问题: • 违背了半保留复制; • DNA复制应在S期,重组应在偶线期,不应同时发生。 (3)不能解释3线和4线双交换。
C. breakage and reunion model(断裂愈合模型) Darlington (1937年)提出 认为重组是复制完成后,两个染色单体断裂并重新连接的结果。 不能解释基因转变(Gene conversion)和极化子(polarity)现象。
D. Holliday model (Holliday模型) (1) Holliday模型 R.Holliday (1964年)提出
(2) Gene conversion(基因转变) ① Gene conversion(基因转变):某一个基因转变为相对的等位基因。
基因转变的发现 1930年,Winkler在真菌发现不规则分离现象; 1949,Lindegren,C.C报道了酵母异离常分离现象; Olive, E1-Ani和Kitani等在粪壳菌(Sodaria fimicola)发现异常分离;
粪生粪壳菌 g+ × g- 5:36:2aberrant 4:4 (0.06%) ( 0.05%) ( 0.008%)
② Types of gene conversion(基因转变的类型) chromatid conversion(6:2) 染色单体转变 half-chromatid conversion (5:3,3:1:1:3) gene conversion 半染色单体转变
+ g × + + 6:2分离 染色单体转变 + + g + g + g 5:3或异常4:4 半染色单体转变 (减数后分离) + g + g
③ Character ofgene conversion(基因转变的特点) A. 基因转变常伴随着重组的发生 例如:A g+×a g-,凡是g+或g-发生转变的,都伴随着A和g之间的重组。 (A、a是一对关于接合型的基因)
A g+ A g+ a g+ a g- A g- A g+ a g- a g- A g+ A g+ a g+ a g+ A g+ A g- a g- a g- 非重组孢子对 非重组孢子对 重组孢子对,基因转变 重组孢子对 重组孢子对,基因转变 重组孢子对,基因转变 非重组孢子对 非重组孢子对 基因转变伴随基因重组
B.基因转变具有极性(Polarity) 同一基因内部的各个突变位点的转变频率常从基因的一端向另一端逐步递减。 具有这种现象的一段DNA称为极化子(polaron) ,一个极化子相当于一个基因。
④ 基因转变的分子机制 Holliday模型中的杂合双链部分校正的时期和方式不同所致。
+/++/+-/- -/- ++++---- 正常4:4 正常校正 G C G T C A A T +/++/++/+-/- ++++++-- 都校正 为+ 6:2 有丝分裂 +/++/-+/- -/- +++-+--- 不校正 3:1:1:3 +/++/++/- -/- +++++--- 一个校正为+ 一个不校正 5:3
不配对碱基由于未及时校正,下一轮DNA复制将产生两个不同的等位基因,通过有丝分裂发生分离,这种现象称减数后分离。不配对碱基由于未及时校正,下一轮DNA复制将产生两个不同的等位基因,通过有丝分裂发生分离,这种现象称减数后分离。 减数后分离导致半染色单体转变。
⑤基因转变与负干涉 当两个基因非常邻近时,特别是在同一个基因的不同突变位点之间,双交换的频率高于预期频率,这种现象称为负干涉。 由基因转变造成。即错配碱基校正结果与该区域双交换结果一致。
校正为 AT(-) A B GT CA a b GC(+) 校正为
+++++--- +/+ +/+ +/- -/- 三型 1 2 3 4 +/++/- +/+ -/- +++-++-- 四型
各种子囊菌中的三型和四型子囊数 三型 四型 四型% Sordaria fimicola 5:3 313 165 35 3:5 204 66 24 Sordaria brevicollis 5:3 83 3 3 3:5 50 1 Ascobolus immersus 5:3 21 0 2 3:5 32 1 三型显著多于四型
E. Meselson-Radding 模型 由 Meselson and Radding 提出。 断裂发生在联会染色体的一条链上; 在 Holliday 模型中,断裂发生在两条非姊妹染色单体的两条链上。
F.双链断裂模型Double-strand-break model 最初由 Jack Szostak, Terry Orr-Weaver, and Rodney Rothstein提出,同一染色单体的两条链断裂引发的重组。
Double-strand-break model of meiotic recombination in the yeast
G. Recombination between circular molecules(环状DNA分子的重组) 一个环旋转180oc 线状二聚体在箭头2、4被切开 单体环在箭头1、3被切开 在箭头1、2或3、4造成相邻切点 环状DNA分子重组过程
校正时切除供体单链核苷酸 ——无重组发生 校正时切除受体单链核苷酸 ——有重组发生 无校正修复 ——子细胞一个重组,一个未重组 有些遗传标记在转化中很少发生校正作用,或是校正切除总是在受体DNA上,转化效率较高,这些遗传标记称为高效率标记(high-efficiency marker)。
有些遗传标记在转化中的校正切除总是倾向发生在供体单链上,转化效率较低,这些遗传标记称为低效率标记(low-efficiency marker)。 校正修复缺陷型体变体中,低效率标记转为高效率标记。
2、Site-specific recombination(位点专一性重组) (1) Site-specific recombination(位点专一性重组):发生在原核生物特殊序列之间的重组过程,又称保守性重组(conservative recombination )。 噬菌体基因组整合到细菌染色体基因组中属此种重组