Download
1 / 22
220 likes | 410 Views
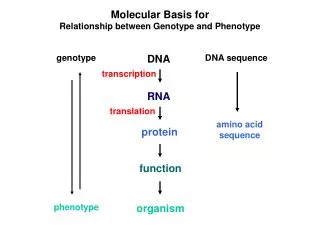
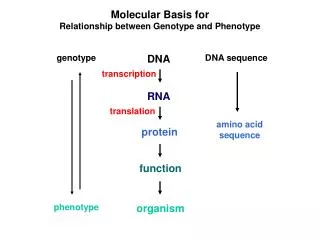
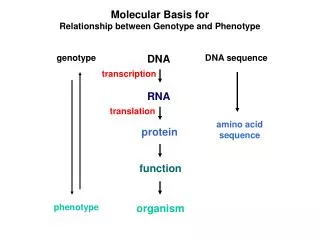
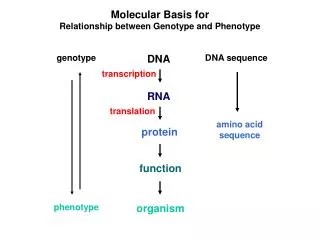
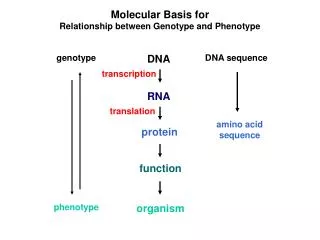
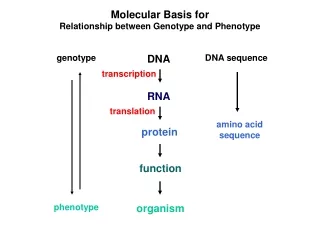
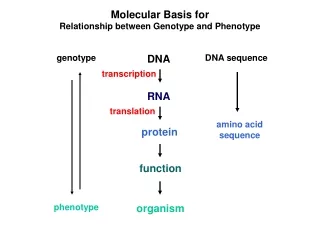
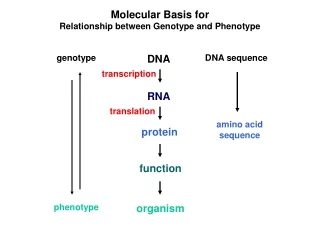
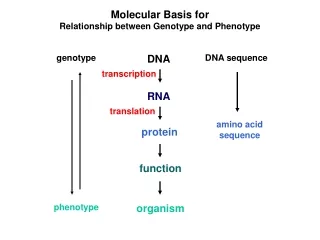
Molecular Basis for Relationship between Genotype and Phenotype. genotype. DNA. DNA sequence. transcription. RNA. translation. amino acid sequence. protein. function. phenotype. organism. Complementarity and Asymmetry in RNA Synthesis.

E N D
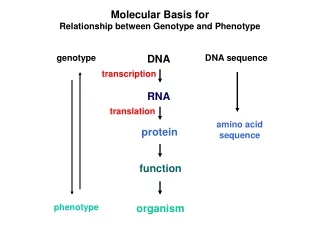
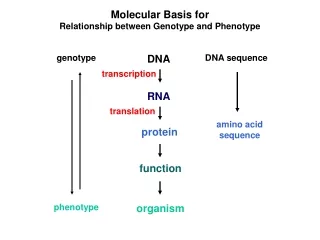
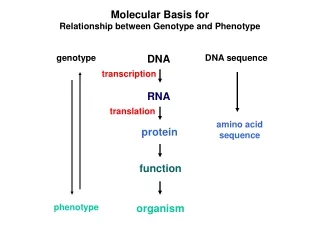
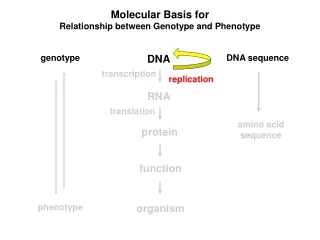
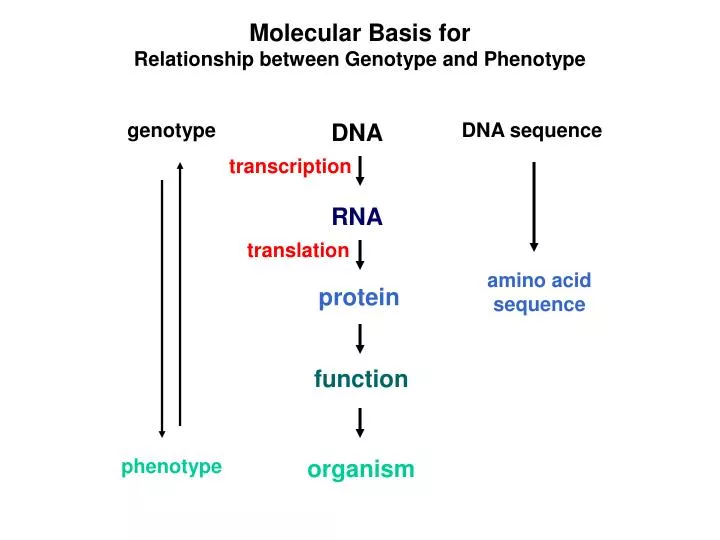
Molecular Basis for Relationship between Genotype and Phenotype genotype DNA DNA sequence transcription RNA translation amino acid sequence protein function phenotype organism
Complementarity and Asymmetry in RNA Synthesis Only one strand of DNA is used as template for RNA synthesis. RNA bases are complementary to bases of template DNA. Template strand is antiparallel to RNA transcript.
RNA Polymerase RNA polymerase in E. coli consists of 4 different subunits (see model below). s recognizes the promoter. Holoenzyme is needed for correct initiation of transcription. RNA polymerase adds ribonucleotides in 5’ to 3’ direction. A single type of RNA polymerase transcribes RNA in prokaryotes.
Promoter Sequences in E. coli Promoters signal transcription in prokaryotes.
Transcription Initiation in Prokaryotes s subunit positions RNA polymerase for correct initiation. Upon initiation of transcription, s subunit dissociates.
Elongation RNA polymerase adds ribonucleotides in 5’ to 3’ direction. RNA polymerase catalyzes the following reaction: DNA Mg++ RNA polymerase NTP + (NMP)n (NMP)n+1 + PPi
Termination Termination of transcription occurs beyond the coding sequence of a gene. This region is 3’ untranslated region (3’ UTR), which is recognized by RNA polymerase.
Termination RNA polymerase recognizes signals for chain termination. (1) Intrinsic: Termination site on template DNA consists of GC-rich sequences followed by A’s. Intra-molecular hydrogen bonding causes formation of hairpin loop. (2) rho factor (hexameric protein) dependent: These termination signals do not produce hairpin loops. rho binds to RNA at rut site. rho pulls RNA away from RNA polymerase. rut site In E. coli, this structure signals release of RNA polymerase, thus terminating transcription.
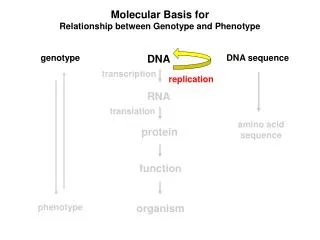
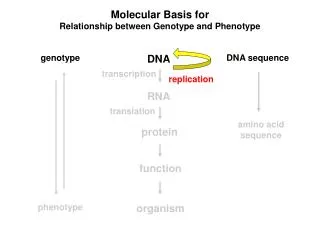
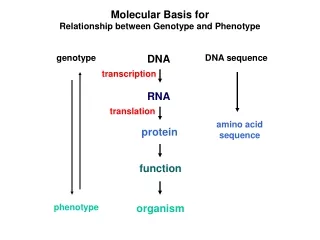
Colinearity of Gene and Protein genotype DNA DNA sequence transcription RNA translation amino acid sequence protein function phenotype organism
Genetic Code Genetic Code is nonoverlapping. A codon (three bases or triplet) encodes an amino acid. Genetic Code is read continuously from a fixed starting point. There is a start codon (AUG). There are three stop (termination) codons. They are often called nonsense codons. Genetic Code is degenerate. Some amino acids are encoded by more than one codon.
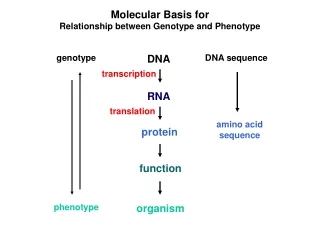
Molecular Basis for Relationship between Genotype and Phenotype genotype DNA DNA sequence transcription RNA translation amino acid sequence protein function phenotype organism
Eukaryotic RNA Three RNA Polymerases RNA Polymerase I II III Synthesis of rRNA (except 5S rRNA) mRNA*, some snRNA tRNA, some snRNA, 5S rRNA * eukaryotic RNA is monocistronic prokaryotic RNA can be polycistronic
Eukaryotic RNA Many proteins must assemble at promoter before transcription. General transcription factors (GTF’s) bind before RNA polymerase II, while other proteins bind after RNA polymerase II binds. Primary transcript (pre-mRNA) must be processed into mature mRNA. 1. Cap at 5’ end (7-methylguanosine) 2. Addition of poly(A) tail 3. Splicing of RNA transcript Chromatin structure affects gene expression (gene transcription) in eukaryotes.
Prokaryotic and Eukaryotic Transcription and Translation Compared
Transcription Initiation in Eukaryotes TATA binding protein (TBP), part of TFIID complex, must bind to promoter before other GTFs and RNA polymerase II can form preinitiation complex (PIC). Phosphorylation of carboxyl tail domain (CTD), the protein tail of b subunit of RNA polymerase II, allows separation of RNA polymerase II from GTFs to start transcription.
Cotranscriptional Processing of RNA State of phosphorylation of CTD determines the type of proteins that can associate with the CTD (thus defining cotranscriptional process). 5’ end of pre-mRNA is capped with 7-methylguanosine. This protects the transcript from degradation; capping is also necessary for translation of mature mRNA.
Cotranscriptional Processing 3’ end of the transcript typically contains AAUAAA or AUUAAA. This sequence is recognized by an enzyme that cleaves the newly synthesized transcript ~20 nucleotides downstream. At the 3’ end, a poly(A) tail consisting of 150 - 200 adenine nucleotides is added. Polyadenylation is another characteristic of transcription in eukaryotes.
Complex Patterns of Eukaryotic RNA Splicing Different mRNA can be produced; different a-tropomyosin can be produced. Alternative splicing is a mechanism for gene regulation. Gene product can be different in different cell types and at different stages of development.
Intron Splicing: Conserved Sequences exons - coding sequences introns - noncoding sequences Small nuclear ribonucleoprotein particles (snRNPs) recognize consensus splice junction sequence of GU/AG. snRNPs are complexes of protein and small nuclear RNA (snRNA). Several snRNPs comprise a spliceosome. Spliceosome directs the removal of introns and joining of exons.
Spliceosome Assembly and Function Spliceosome interacts with CTD and attaches to pre-mRNA. snRNAs in spliceosomes direct alignment of the splice sites. One end of conserved sequence attaches to conserved adenine in the intron. The “lariat” is released and adjacent exons are joined.
Self-Splicing Reaction RNA molecules can act somewhat like enzymes (ribozymes). In the protozoan Tetrahymena, the primary transcript of an rRNA can excise a 413-nucleotide intron from itself. These self-splicing introns are an example of RNA that can catalyze a reaction.