Download
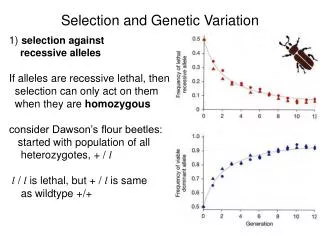
1 / 35
350 likes | 481 Views
Non-stationary population genetic models with selection: Theory and Inference. Scott Williamson and Carlos Bustamante. Cornell University. Inferring natural selection from samples. Statistical tests of the neutral theory (lots) Methods for detecting selective sweeps (lots)

E N D
Non-stationary population genetic models with selection:Theory and Inference Scott Williamson and Carlos Bustamante Cornell University
Inferring natural selection from samples • Statistical tests of the neutral theory (lots) • Methods for detecting selective sweeps (lots) • Parametric inference: estimating selection parameters, etc. • Quantification of selective constraint, deleterious mutation
The demography problem • Many existing methods assume random mating, constant population size • These assumptions don’t apply in most natural populations • The effect of demography can mimic the effect of natural selection
Natural selection and population growth • Inferring selection from the frequency spectrum while correcting for demography • The McDonald-Kreitman test: does recent population growth cause you to misidentify negative selection as adaptive evolution?
7019 4424 4961 5286 1972 2188 3529 163 975 A G G C T T A A A 1 A T G C T C G A A 2 G T G T T C A C G 3 A G G C T C A A G 4 A G A C C C G A A 5 1 2 1 1 1 4 2 1 3 The frequency spectrum: an example The frequency spectrum Site Sequence Count Frequency class: Frequency class Ancestral Derived
Natural selection and the frequency spectrum Equilibrium neutral and positively selected frequency spectra Neutral 2Ns=2 Count Frequency class
Natural selection and the frequency spectrum Equilibrium neutral and negatively selected frequency spectra Neutral 2Ns=-2 Count Frequency class
Natural selection vs. demography Non-stationary neutral and equilibrium selected frequency spectra Population growth, neutral Equilibrium, 2Ns=-2 Count Frequency class
How do we distinguish selection from demography? • McDonald-Kreitman approach: • Use a priori information to classify changes as “neutral” (e.g. synonymous, non-coding) or “potentially selected” (e.g. non-synonymous) • Putatively neutral changes are treated as a standard for patterns of neutral evolution in a particular sample • Potentially selected sites are compared to the neutral standard Can we develop a neutral standard for the frequency spectrum?
Comparing frequency spectra for different classes of mutation Observed frequency spectra • This talk: • Likelihood ratio test of neutrality at potentially selected sites, using information from the neutral sites • Biologically meaningful measure of the difference between the two spectra Putatively neutral Potentially selected Count Frequency class
Comparing frequency spectra for different classes of mutation Observed frequency spectra A model-based approach: • Fit a neutral demographic model to estimate demographic parameters Putatively neutral Potentially selected Count • Given those parameter estimates, fit a selective demographic model to estimate selection parameters, test hypotheses Frequency class
Comparing frequency spectra for different classes of mutation Observed frequency spectra • Requirements: • Demographic model • Frequency spectrum predictions from the model under neutrality • Frequency spectrum predictions from the model subject to natural selection Putatively neutral Potentially selected Count Frequency class
Theory: population growth model 2-epoch model NC Population size NA =NA/NC time now Model parameters: ,
Theory: predicting the frequency spectrum Definitions: xi Number of sites in frequency class i f(q,t;) Distribution of allele frequency, q, at time t n Sample size Predictions:
Theory: the distribution of allele frequency Poisson Random Field approach (Sawyer and Hartl 1992): • Use single-locus diffusion theory to predict the distribution of allele-frequency • If sites are independent (i.e. in linkage equilibrium) and identically distributed, then the single-locus theory applies across sites To get f, we need to solve the diffusion equation:
Theory: time-dependent solution, neutral case The forward equation under neutrality: Kimura’s (1964) solution, given some initial allele frequency, p:
Theory: time-dependent solution, neutral case Applying Kimura’s solution to the 2-epoch model: ancestral mutations Kimura’s (1964) solution, given some initial allele frequency, p: Distribution of allele frequency:
Theory: time-dependent solution, neutral case Expected frequency spectrum after a change in population size (=0.01) 0.8 0.6 P(i,n;,0.01) 0.4 0.2 1 2 3 4 5 6 7 8 9 frequency class
Theory: time-dependent solution, neutral case Multinomial likelihood: Maximum likelihood estimates of and Likelihood ratio test of population growth
Comparing frequency spectra for different classes of mutation Observed frequency spectra • Requirements: • Demographic model • Frequency spectrum predictions from the model under neutrality • Frequency spectrum predictions from the model subject to natural selection Putatively neutral Potentially selected Count Frequency class
Theory: time-dependent solution, selected case The forward equation with selection: where =2NCs Initial condition:
Theory: time-dependent solution, selected case • Numerically solve the forward equation using the Crank-Nicolson finite differencing scheme • Use this approximation of f to evaluate the likelihood function: • Fix and to their MLEs from the neutral data • Optimize the likelihood for . Likelihood ratio test of neutrality:
Theory: time-dependent solution, selected case How can we be sure that the numerical solution actually works? • Von Neumann stability analysis: solution is unconditionally stable • Numerical solution converges to the stationary distribution after ~4NC generations • Comparison with time-dependent neutral predictions: Kimura, Crank, and Nicolson all agree with each other
Human Polymorphism Data • From Stephens et al. (2001) • 80 individuals, geographically diverse ancestry • 313 genes, 720 kb sequenced • ~3000 SNPs (72% non-coding, 13% synonymous, 15% non-synonymous)
Results for non-synonymous changes, categorized by Grantham’s distance
Ongoing work and future directions • Simulate, simulate, simulate • How robust is the method to different types of demographic forces? • How does linkage among some sites affect the analysis? • How does estimation error affect the LRTs? • Numerical solution for different demographic scenarios (e.g. bottleneck, population structure) • Variable selective effects among new mutations
The McDonald-Kreitman test Sn Number of non-synonymous segregating sites Dn Number of non-synonymous fixed differences Ss Number of synonymous segregating sites Ds Number of synonymous fixed differences Adaptive evolution Negative selection Extensions: Sawyer and Hartl (1992), Rand and Kann (1996), Smith and Eyre-Walker (2002), Bustamante et al. (2002), others
Demography and the McDonald-Kreitman test • Robust to different demographic scenarios because it implicitly conditions on the underlying genealogy (see Nielsen 2001) • However, under some demographic scenarios it’s possible to misidentify the type of selection • Weak negative selection with population growth • When the population size is small, non-synonymous deleterious mutations might be fixed by drift • Once the population size becomes large, the level of non-synonymous polymorphism would be reduced (relative to the level of synonymous polymorphism)
Demography and the McDonald-Kreitman test • Over what range of parameter values might you misidentify negative selection as adaptive evolution? • How large is the effect? Eyre-Walker (2002): • Addressed these questions, finding that recent population growth or bottlenecks can cause you to misidentify negative selection • Assumed that levels of polymorphism and fixation rates changed instantaneously with population size
Demography and the McDonald-Kreitman test where tdiv is the divergence time, measured in 2NC generations
Demography and the McDonald-Kreitman test =0.1, tdiv=10 10 10 =0.1, tdiv=4 1 1 0.1 1 0.1 1 0.01 0.01 Expected Neutrality Index (NI) =1, tdiv=4 =1, tdiv=10 10 10 1 1 0.1 1 0.1 1 0.01 0.01 (=NA/NC)
Demography and the McDonald-Kreitman test: Preliminary results • It is possible to misidentify negative selection for some parameter combinations • But…the parameter range over which this is true is probably smaller than previously thought, as is the magnitude of the effect
Summary • Model-based approach to correcting for demography while inferring selection • Evidence for very recent population growth in humans • Reasonable estimates of selection parameters for classes of non-synonymous changes • McDonald-Kreitman test: negative selection + population growth problem not as severe as previously thought • Numerical methods for solving the diffusion are fast, accurate, and fun!
Collaborator:Carlos Bustamante Acknowledgements Data:Genaissance Pharmaceuticals Helpful discussions:Bret Payseur, Rasmus Nielsen, Matt Dimmic, Jim Crow, Hiroshi Akashi, Graham Coop