Download

1 / 37
370 likes | 587 Views
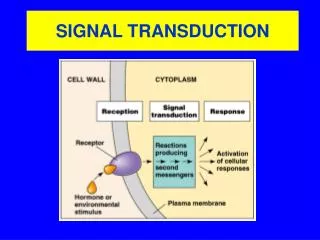
Photo-transduction. and related mathematical problems. D. Holcman, Weizmann Institute of Science. Retinal organization. Retina connection. Cone > Bipolar cell > Ganglion cell Rod > Bipolar cell > Amacrine cell > Ganglion cell. Photo-response cone/rod. Actual state of art.

E N D
Photo-transduction and related mathematical problems D. Holcman, Weizmann Institute of Science
Retina connection • Cone > Bipolar cell > Ganglion cell • Rod > Bipolar cell > Amacrine cell > Ganglion cell
Actual state of art • Initial phase of the transduction known • The global recovery is still missing • Difference of the two photoreceptors? • How signal propagate from the outer-segment to the synapse? • How the synapse is modulated?
Steps of Photo-transduction • 1-Arrival of a photon: RhRh* • 2-Amplification from Rh*…PDE* a single Rh^* activates 300 PDE • 3-Destruction of cGMP messenger • 4-Channels closed • 5-hyper-polarization of the cell • 6-Transmission like a wave capacitance to the Inner-Segment • 7-Release of neurotransmitters
Order of magnitude Number per compartment of • cGMP: 60 to 200 • Channels 200 to 300 • Open channels in dark= 6 • Activated PDE=1 • Free calcium =5 Photon close channels: Can closing 6 enough to generate a signal?
Longitudinal propagation of a signal • cGMP holes propagate to close many channels: how much? • Compute the propagation of the depleted area
A theory of longitudinal diffusion at a molecular level F electrostatic forces w noise Particle motion in the Outer Segment The pdf satisfies the following equations within the outer segment F=0. where and m mass of the molecule g viscosity coefficient T absolute temperature k Boltzmann constant
Longitudinal diffusion in rod outer segments • Method: projection 3D1D Conclusion: standard linear diffusion
Longitudinal diffusion in cone outer segments • Method: projection 3D1D d diameter of disc connecting two adjacent compartments D Diffusion constant d min diameter at the tip CONCLUSION 1-the diffusion coefficient is not a constant value, but change with longitudinal position 2-No explicit solution (WKB asymptotic)
Spread of excitation cGMP =messenger that open channels 1-Compare spread of cGMP in rod/cone 2- Characterize the spread at time to peak tp of the photo-response
Species COS structure cGMP diffusion Species ROS structure cGMP diffusion Length (mm) Base radius (mm) Tip radius (mm) d (mm) Dl(base) (mm2/sec) Dl (tip) (mm2/sec) Dl (at L/2) (mm2/sec) lcon (at L/2) (mm) length (mm) diameter (mm) No. incisures Daq (mm2/sec) Dl (experiment) (mm2/sec) lrod (mm) Striped bass, single cone 15 3.1 1.2 20.324 2.7 17.9 5.6 40.79 Tiger salamander 125.3 12.3 218 500 330-60 21-11 18.5 84.7 Tiger salamander, single cone 8.5 2.5 1.1 30.314 3.9 20.0 7.6 50.99 7Striped bass 40 1.6 1 41.6 73.8 Human, peripheral retina1 7 1.5 0.75 30.244 6.6 25.8 11.6 60.68 4 Human, peripheral retina 12 1.5 1 44.3 93.0 5Guinea pig 5 1.4 1 47.3 6Rat 25 1.7 1 39.3 Dl (theory) (mm2/sec) Comparison across species of spread of excitation • our data, n=11
Conclusion on the longitudinal diffusion 1-Spread of Excitation depends on the geometry only but not on the size. 2-Geometry alone determines the longitudinal diffusion 2-Spread of excitation is similar across species for Cones and Rods D. Holcman et al. Biophysical Journal, 2004l
Access to all global variable • Membrane potential V(t) Total Calcium and cGMP
Conclusion • Presented here a global model • Simulate photo-response from 1 to many • Adaptation is not included
fluctuation of the membrane potential G. Field. F.Rieke, Neuron 2002
Sources of Noise • Definition: fluctuation of the membrane potential Causes • Thermal activation of Rhodopsin • Local binding and unbinding of CGMP + Push-pull mechanism (swimming noise) • PDE activity as a source of the noise in chemical reactions: Push-Pull noise
Swimming noise • Fluctuation of the number of open channels due the stochastic binding and unbinding.
Swimming noise • Number of open channels (experimentally=6) • Variance= compute? Model Rules: • cGMP bind and unbind to the channels, diffuse inside a compartment • When a channel is gated, no other cGMP can bind. • cGMP stays bound during a given time.
Swimming noise = number of unbound particles at time = number of free sites in volume at time = number of unbound binding sites at time = number of bound particles at time . = initial density of substrate The joint probability of a trajectory and the number of bound sites in the volume
Fokker-Planck Equation for the joint pdf • P(x,S,t)= proba to find a cGMP at position x at time t and S(0 or 1) channel are bound at position x • Time evolution equation J=flux, K1 redined forward binding, k-1 backward rate
Steady state Parabolic variance
Push-Pull mechanism Fact: cGMP is regulated by 1 PDE* and another moleculetotal number of cGMP fluctuate Continuum model Steady state variance can be computed from the same analysis
Conclusion • Simulation is needed • Include cooperativity effect (up to 4 cGMP can be bound to a single channel) • Derive the fluctuation of the number of open channels and the characteristic time • Derive a Master equation to compute mean and variance of the cGMP due to the Push-pull.
Where we stand:Push-Pull noise, low frequency • Molecular difference of the steady state noise (RGS9PDE*) • Description of the noise: a problem of Mean First Passage Time in chemical reactions
Simplifies Model • cGMP fluctuation due to the push-pull (no diffusion) N* colored noise= fluctuation of independent PDE K, a,b, sigma, gamma constant W=Brownian Characterization of the fluctuationin CGMP= Find the MFPT of c to a threshold as a function of the parameter
Mean First Passage Time • Attractor (c,N*)= • p not the same for cones and rods Kind of Smoluchowski limit
Fokker Planck Operator Find P0