Download
1 / 27
280 likes | 462 Views
Diffusible morphogen. Neuroinduction. Ant. Post. Endoderm and Mesoderm involute with gastrulation: Induction of the Neural Plate from Neuroectoderm, by the underlying , closely apposed Mesoderm. Hilde Mangold and Hans Spemann. Key experiments performed in 1921-1923 at the

E N D
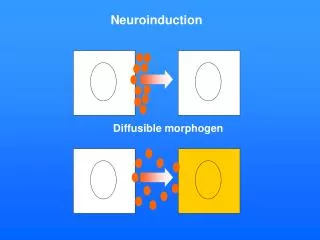
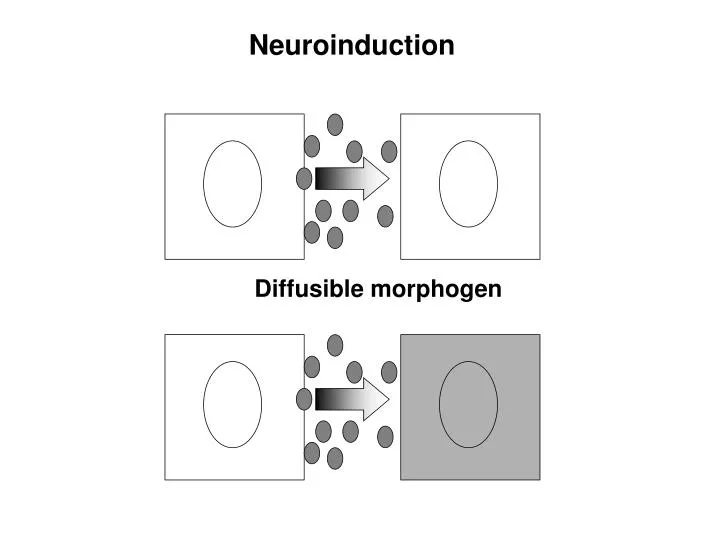
Diffusible morphogen Neuroinduction
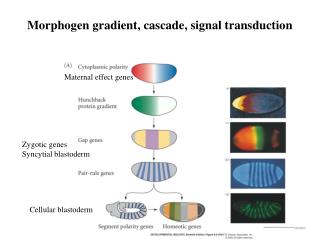
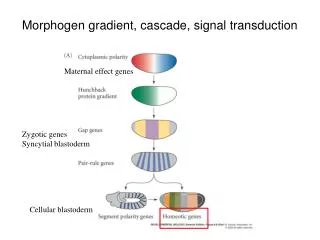
Ant Post Endoderm and Mesoderm involute with gastrulation: Induction of the Neural Plate from Neuroectoderm, by the underlying, closely apposed Mesoderm.
Hilde Mangold and Hans Spemann • Key experiments performed in 1921-1923 at the • University of Freiburg, Germany. • Hilde Mangold was a 24 year old graduate • student when she performed these experiments. • She died tragically in an accidental alcohol heater • explosion. • Hans Spemann was awarded the Nobel Prize in 1935.
Hilde Mangold and Hans Spemann Experiments (1924)
! Explant experiments with Animal Caps from AmphibianBlastula: Puzzling results…
+ Candidate Neuroinducing Factors ? (Intact) Isolating inducing factors that promote Neuronal differentiation; The “Sigma catalog” experiments further confusion…
Model 1: +”Epidermal factor” Epidermis Presumptive Neuroectoderm Neurons +”Neuronal factor” Model 2: Epidermis Presumptive Neuroectoderm (“default”) Neurons +”Neuronal factor” Model 3: +”Epidermal factor” Epidermis Presumptive Neuroectoderm Neurons (“default”) Models for Neural Induction
TGF-b Proteins Signal Through Heterodimeric Receptors and Smad Transcription Factors
A Dominant-Negative Receptor Subunit Blocks Activation of the Signaling Pathway (Hemmati-Brivanlou and Melton, 1992)
(+Dominant-Negative Type II Receptor cRNA) TFG-b Signaling Blocked by expression of Dom-Neg Type II Receptor Subunit Animal Cap (Intact) Animal Cap (Intact) + TGF-b Signaling + TGF-b Signaling (Intact) Blocking TGF-b signaling by a dominant-negative receptor causes isolated Neuroectoderm to become Neuronal
BMP-4 (TGF-b) Signaling Results in “NeuralEpidermal Induction” TGF-b: Transforming GrowthFactor - b BMP-4: BoneMorphogenic Protein - 4
Model 1: Epidermis Presumptive Neuroectoderm Neurons +”Neuronal factor” Model 2: Epidermis Presumptive Neuroectoderm (“default”) Neurons +”Neuronal factor” Model 3: Epidermis Presumptive Neuroectoderm Neurons (“default”) Models for Neural Induction +”Epidermal factor” +BMP-4
+ BMP-4 [BMP-4] BMP-4 (Secreted by Neuroectodermal Cells) Inhibits Neuronal Fate and Promotes Epidermal Fate. Tissue Dissociation dilutes BMP-4 activity (Endogenous BMP-4 Diluted)
Dispersed caps Intact caps Keratin (epidermal marker) NCAM (neuronal marker) (Wilson and Hemmati-Brivanlou, 1995) Recombinant BMP-4 promotes epidermal fate and inhibits neuronal fate
(Fainsod, et al., 1995) BMP-4 mRNA is expressed in presumptive ectoderm Blastopore
Are there native anatgonists of BMP-4? Secreted from underlying mesoderm? Yes… chordin / noggin / follistatin. And they are enriched in the Spemann-Mangold Organizer!
Noggin cRNA injections rescue ventralized embryos Chordin expresses in mesoderm +Noggin injection 1pg 10pg (Sasai, et al., 1995) 100pg (Smith and Harland, 1992)
Differential Substractive Screen yields Chordin, a BMP-4 antagonist (1994) (Sasai and DeRobertis. 1994)
Functional Expression Cloning yields noggin, a BMP-4 anatagonist (1992) (Smith and Harlan, 1992)
(Migrates into gel) (Stays in loading well) Mechanism: Chordin / noggin / follistatin anatagonize BMP-4 activity by directly binding and inactivating BMP-4
noggin Receptor (Type-II) Receptor (Type-I) BMP-7 Crystal structure of Noggin-BMP complex confirms biochemical/functional studies and identifies binding sites Binding Sites (Groppe, et al., 2002)
TGF-b proteins signal through heterodimeric receptors and Smad transcription factors
Vertebrates Drosophila Ligand Receptor Antagonist Transcription Factor BMP-4 Type I Type II Type III noggin chordin follistatin Smad1 Smad2 Smad3 Smad4 Smad5 decapentaplegic (dpp) punt thick veins (tkv), saxophone (sax) Short-gastrulation (sog) Mothers against decapentaplegic (MAD) Medea The mechanism for neural induction is evolutionarily conserved between vertebrates and invertebrates
BMP-4 is only one member of the large evolutionarily conserved TGF-b gene family, which mediates many different tissue inductive events.
Neurogenesis: Inductive Mechanisms 1. Neuroectodermal cells choose either a neuronal or epidermal cell fate. 2. Interactions between mesoderm and neuroectoderm induce neuroectoderm to adopt the neural fate. 3. Induction acts through signaling by a secreted protein, Bone Morphogenic Protein-4 (BMP-4), made by neuroectodermal cells. 4. BMP-4 inhibits neuralization and promotes the epidermal fate in neighboring cells. 5. Mesodermal cells secrete proteins (Chordin, Noggin, Follistatin) which directly bind and antagonizes BMP-4 activity. 6. Neuroectodermal cells become neurons by suppression of BMP-4 activity by secreted antagonists from underlying mesodermal cells.
Neurogenesis: Inductive Mechanisms (cont.) 7. The “default” state of neuroectodermal cells is neuronal. 8. This inductive mechanism is conserved between vertebrates and invertebrates. 9. BMP-4 is a member of the Transforming Growth Factor (TGF-b) family of signaling molecules. Similar signaling events maybe selectively and focally re-employed later in the nervous system to mediate fine tuning at late developmental stages and adult plasticity.