Download

1 / 69
710 likes | 737 Views
Enzymes : Mechanism and Catalysis. Enzymes DO NOT change the equilibrium constant of a reaction Enzymes DO NOT alter the amount of energy consumed or liberated in the reaction ( standard free energy change, G° ) Enzymes DO increase the rate of reactions that are otherwise possible

E N D
Enzymes : Mechanism and Catalysis
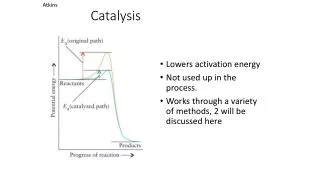
Enzymes DO NOT change the equilibrium constant of a reaction • Enzymes DO NOT alter the amount of energy consumed or liberated in the reaction (standard free energy change, G°) • Enzymes DO increase the rate of reactions that are otherwise possible • Enzymes DO decrease the activation energy of a reaction (G°‡)
Enzymes DO increase the rate of reactions that are otherwise possible • Enzymes DO decrease the activation energy of a reaction (G°‡) The classic way that an enzyme increases the rate of a bimolecular reaction is to use binding energy to simply bring the two reactants in close proximity. In order for a reaction to take place between two molecules, the molecules must first find each other. This is why the rate of a reaction is dependent upon the concentrations of the reactants, since there is a higher probability that two molecules will collide at high concentrations. The enzyme organizes the reaction at the active site, thereby reducing the cost in terms of ENTROPY.
How do enzymes catalyze biochemical reactions? • involves basic principles of organic chemistry • What functional groups can be involved in catalysis? • almost all alpha amino and carboxyl groups are tied up in peptide bonds • R groups are involved in catalysis • asp, glu • his, lys • ser, cys, tyr • catalysis occurs when substrate is immobilized near these residues at the active site
General Acid-Base Catalysis • General acid-base catalysis is involved in a majority of enzymatic reactions. General acid–base catalysis needs to be distinguished from specific acid–base catalysis. • In General acid–base catalysis, the buffer aids in stabilizing the transition state via donation or removal of a proton. Therefore, the rate of the reaction is dependent on the buffer concentration, as well as the appropriate protonation state.
General Base Catalysis II:Ester Hydrolysis The hydrolysis of esters proceeds readily under in the presence of hydroxide. It is base catalyzed. However, the rate of hydrolysis is also dependent on imidazole buffer concentration. Imidazole can accept a proton from water in the transiton state in order to generate the better nucleophile, hydroxide. It can also re-donate the proton to the paranitrophenylacetate in order to generate a good leaving group.
General Acid Catalysis :Ester Hydrolysis Electrostatic interactions are much stronger in organic solvents than in water due to the dielectric constant of the medium. The interior of enzymes have dielectric constants that are similar to hexane or chloroform
Catalysis by Metal Ions Catalysis I:Ester Hydrolysis Metal ions that are bound to the protein (prosthetic groups or cofactors) can also aid in catalysis. In this case, Zinc is acting as a Lewis acid. It coordinates to the non-bonding electrons of the carbonyl, inducing charge separation, and making the carbon more electrophilic, or more susceptible to nucleophilic attack.
Catalysis by Metal Ions Catalysis II:Ester Hydrolysis Metal ions can also function to make potential nucleophiles (such as water) more nucleophilic. For example, the pka of water drops from 15.7 to 6-7 when it is coordinated to Zinc or Cobalt. The hydroxide ion is 4 orders of magnitude more nucleophilic than is water.
Enzymes physically interact with their substrates to effect catalysis • E + S ES ES* EP E + P • where… • E = enzyme • S = substrate • ES = enzyme/substrate complex • ES* = enzyme/transition state complex • P = product • EP = enzyme/product complex • Enzyme and substrate combine to form a complex • Complex goes through a transition state (ES*) • bound substance is neither substrate nor product • A complex of the enzyme and the product is formed • The enzyme and product separate • All of these steps are governed byequilibria
Substrates bind to the enzyme’s active site • pocket in the enzyme • Substrates bind in active site by • hydrogen bonding • hydrophobic interactions • ionic interactions
Enzyme/Substrate Interactions • Lock and key model • substrate (key) fits into a perfectly shaped space in the enzyme (lock) • Induced fit model • substrate fits into a space in the enzyme, causing the enzyme to change conformation • change in protein conformation leads to an exact fit of substrate with enzyme • Following catalysis, the product(s) no longer fits the active site and is released Enzymes and Enzyme Kinetics
Strain and Distortion model • The binding of the substrate results in the distortion of the substrate in a way that makes the chemical reaction easier.
Enzyme Kinetics • The rate of the reaction catalyzed by enzyme E • A + B P • is defined as • -D[A] or -D[B] or D[P] • Dt Dt Dt • Enzyme activity can be assayed in many ways • disappearance of substrate • appearance of product • continuous assay • end point assay • For example, you could measure • appearance of colored product made from an uncolored substrate • appearance of a UV absorbent product made from a non-UV-absorbent substrate • appearance of radioactive product made from radioactive substrate Enzymes and Enzyme Kinetics
Higher temperature generally causes more collisions among the molecules and therefore increases the rate of a reaction. More collisions increase the likelihood that substrate will collide with the active site of the enzyme, thus increasing the rate of an enzyme-catalyzed reaction. Each enzyme has an optimal pH. A change in pH can alter the ionization of the R groups of the amino acids. When the charges on the amino acids change, hydrogen bonding within the protein molecule change and the molecule changes shape. The new shape may not be effective. The diagram shows that pepsin functions best in an acid environment. This makes sense because pepsin is an enzyme that is normally found in the stomach where the pH is low due to the presence of hydrochloric acid. Trypsin is found in the duodenum, and therefore, its optimum pH is in the neutral range to match the pH of the duodenum.
At lower concentrations, the active sites on most of the enzyme molecules are not filled because there is not much substrate. Higher concentrations cause more collisions between the molecules. With more molecules and collisions, enzymes are more likely to encounter molecules of reactant. The maximum velocity of a reaction is reached when the active sites are almost continuously filled. Increased substrate concentration after this point will not increase the rate. Reaction rate therefore increases as substrate concentration is increased but it levels off. If there is insufficient enzyme present, the reaction will not proceed as fast as it otherwise would because there is not enough enzyme for all of the reactant molecules. As the amount of enzyme is increased, the rate of reaction increases. If there are more enzyme molecules than are needed, adding additional enzyme will not increase the rate. Reaction rate therefore increases as enzyme concentration increases but then it levels off.
The velocity (V) of an enzyme-catalyzed reaction is dependent upon the substrate concentration [S] • A plot of V vs [S] is often hyperbolic (Michaelis-Menten plot) Enzymes and Enzyme Kinetics
The Michaelis-Menten equation describes the kinetic behavior of many enzymes • This equation is based upon the following reaction: • S P • k1 k2 • E + S ES E + P • k-1 • V = Vmax [S] • KM + [S] • the reverse reaction (P S) is not considered because the equation describes initial rates when [P] is near zero Enzymes and Enzyme Kinetics
V = Vmax [S] • KM + [S] • V is the reaction rate (velocity) at a substrate concentration [S] • Vmax is the maximum rate that can be observed in the reaction • substrate is present in excess • enzyme can be saturated (zero order reaction) • KM is the Michaelis constant • a constant that is related to the affinity of the enzyme for the substrate • units are in terms of concentration • KM = k-1 + k2 • k1 Enzymes and Enzyme Kinetics
Enzyme (E) Substrate (S) K_M (in M) k_{cat} (in s^{-1}) k_{cat}/K_M (in M^{-1}s^{-1}) Acetylycholine esterase Acetylcholine 9.5 x 10^{-5} 1.4 x 10^4 1.5 x 10^6 Carbonic Anhydrase CO_2 0.012 1.0 x 10^6 8.3 x 10^7 Carbonic Anhydrase HCO_3- 0.026 4.0 x 10^5 1.5 x 10^7 Catalase H_2O_2 0.025 1.0 x 10^7 4.0 x 10^8 Fumerase Fumerate 5.0 x 10^{-6} 800 1.6 x 10^8 Fumerase Malate 2.5 x 10^{-5} 900 3.6 x 10^7 Urease Urea 0.025 1.0 x 10^4 4.0 x 10^5
KM is also the substrate concentration at which the enzyme operates at one half of its maximum velocity • if [S] = KM • V = Vmax [S] • 2[S] • V = Vmax • 2 Enzymes and Enzyme Kinetics
To determine KM and Vmax, decide on a number of different [S] values, and measure V at each concentration (hold [E] constant) Enzymes and Enzyme Kinetics
Michaelis-Menten plot is not useful for estimating KM and Vmax • it is better to transform the Michaelis-Menten equation to a linear form • actual values for KM and Vmax determined from graph • double reciprocal plot or a Lineweaver-Burk plot Enzymes and Enzyme Kinetics
By taking the inverse of the Michaelis-Menten equation • same form as y = mx + b • plot is y vs x • y is 1/V • x is 1/[S] • KM/Vmaxis slope • y intercept is 1/Vmax • x intercept is -1/ KM V = Vmax [S] KM + [S] 1 = KM + [S] V Vmax [S] Vmax [S] 1 = KM . 1 + 1 V Vmax [S] Vmax Enzymes and Enzyme Kinetics
Enzyme Inhibition • Certain compounds inhibit enzymes • decrease the rates of their catalysis • inhibition can be reversible or irreversible • 3 types of reversibleinhibitors • competitive inhibitors • non-competitive inhibitors • un-competitive inhibitors • Irreversible inhibition • suicide inhibitors • the various types of inhibitors can be distinguished by the kinetics of their inhibition Enzymes and Enzyme Kinetics
Competitive inhibition • inhibitor mimics substrate • fits into active site • malonate is a competitive inhibitor of succinate dehydrogenase Enzymes and Enzyme Kinetics
Competitive inhibitors can be identified by the kinetics of their inhibition • In the presence of a competitive inhibitor • KM increases • Vmax stays the same • The effects of competitive inhibition can be overcome by increasing [S] Enzymes and Enzyme Kinetics
Non-competitive inhibition • inhibitor binds to a site other than the active site • Non-competitive inhibitors can be identified by the kinetics of their inhibition • In the presence of a non-competitive inhibitor • KM stays the same • Vmax decreases • The effects of non-competitive inhibition cannot be overcome by increasing [S] Enzymes and Enzyme Kinetics
Un-competitive inhibition • inhibitor binds to a site other than the active site, but only when substrate is bound • Un-competitive inhibitors can be identified by the kinetics of their inhibition • In the presence of an un-competitive inhibitor • KM decreases • Vmax decreases • The effects of un-competitive inhibition cannot be overcome by increasing [S] Enzymes and Enzyme Kinetics
Irreversible inhibition • enzyme is covalentlymodified after interaction with inhibitor • derivatized enzyme is no longer a catalyst • Organofluorophosphates used as insecticides and nerve gases • irreversible inhibitors of acetylcholinesterase • form covalent product with active site serine residue • enzyme no longer functional Enzymes and Enzyme Kinetics
When chymotrypsin is treated with DIPF • only ser 195 reacts is derivatized • other ser residues are not labeled • ser 195 is in the enzyme’s active site • Why is only ser 195 labeled? • adjacent amino acid residues in active site make ser 195 more reactive Enzymes and Enzyme Kinetics
The Acyl Enzyme Intermediate Diisopropylflurophosphate is an inhibitor of chymotrypsin. It diffuses into the active, wherein a nucleophilic amino acid attacks the phosphate, releasing fluoride anion. This results in a covalent bond between the nucleophile and the inhibitor. It inhibits the reaction because it blocks entry of normal substrates. The enzyme-inhibitor adduct is very stable. Upon hydrolysis of the protein (6 N HCl, 110°C) and amino acid analysis on the hydrolysate, a novel amino acid was isolated. It was the diisopropylphosphoryl derivative of serine.
Coenzymes and Prosthetic Groups • some enzymes employ coenzymes and prosthetic groups at their active sites • used for reactions that amino acid R groups can’t perform • coenzymes • metals or small organic molecules • not covalently bound to protein • often function as co-substrates • precursors are often vitamins • prosthetic groups • small organic molecules • covalently linked to protein Enzymes and Enzyme Kinetics
Enzyme Regulation : 1. Control of Enzyme Activity Level A. Noncovalent modifiers cause conformational change between less active and more active states of the enzyme. B. Covalent Modification causes interconversion between inactive and active forms of the enzyme. 2. Control the Amount of the Enzyme A. Isozymes - forms of the enzyme which differ in properties but catalyze the same reaction. For example, enzyme forms which differ in Vmax and/or Km. The isozymes can be forms found in different tissues and organs of an animal or for any eukaryotic organism, isozymes can be located in different parts of the cell. For example, different isozymes of lactate dehydrogenase are found in muscle and liver. Malate dehydrogenase occurs in different forms in the cytoplasm and the soluble matrix phase of the mitochondria. B. Biosynthesis of the enzyme protein can be controlled at the level of the gene via regulation of transcription (ie synthesis of the enzyme's mRNA). This is more of a molecular biologic type of regulation and involves molecules which bind to DNA and influence gene expression. This type of control where the amount of the enzyme is governed can also be done after the mRNA is made, but this is quite rare. In this mechanism, the mRNA is prevented from being translated and since mRNA is rather unstable, it is degraded before it is effectively used by the ribosomes to make the protein.
Allosteric Regulation Control of Enzyme Activity by Non-Covalent Modifiers is usually called allosteric regulation since the modifier binds to the enzyme at a site other than the active site but alters the shape of the active site. Allosteric is a word derived from two Greek words: 'allo' meaning other and 'steric' meaning place or site; so allosteric means other site and an 'allosteric enzyme' is one with two binding sites - one for the substrate and one for the allosteric modifier molecule, which is not changed by the enzyme so it is not a substrate. The molecule binding at the allosteric site is not called an inhibitor because it does not necessarily have to cause inhibition - so they are called modifiers. A negative allosteric modifier will cause the enzyme to have less activity, while a positive allosteric modifier will cause the enzyme to be more active. In order for allosteric regulation to work, the enzyme must be multimeric (ie. a dimer, trimer, tetramer etc.). The concept is easily illustrated using a dimer as the model system, but it applies equally well to higher order multimers such as trimers and tetramers, etc.
Cooperativity Enzyme can bind two substrates molecules at different binding sites. k1 k2 S + E C1 P + E k-1 k3 k4 S + C1 C2 P + E k-3 or S S E C1 C2 S S P P E E