Download

1 / 32
330 likes | 578 Views
Lateral gene transfer in prokaryotic genomes. Uri Gophna Dept. of Molecular Microbiology and Biotechnology, TAU. Lateral (or Horizontal) Gene Transfer : A process in which an organism transfers genetic material (i.e. DNA ) to another cell that is not its offspring. Outline of the talk.

E N D
Lateral gene transfer in prokaryotic genomes Uri Gophna Dept. of Molecular Microbiology and Biotechnology, TAU
Lateral (or Horizontal) Gene Transfer : A process in which an organism transfers genetic material (i.e. DNA) to another cell that is not its offspring.
Outline of the talk • Molecular mechanisms of lateral gene transfer (LGT) in microorganisms. • Genomic evidence for the importance of LGT. • LGT and the tree of life. • LGT detection methods. • A final note – which transferred genes are fixed in populations?
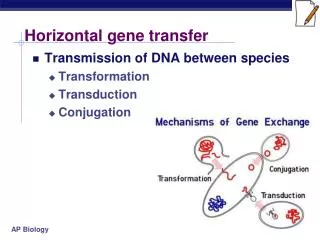
Molecular mechanisms of lateral gene transfer (LGT) • Uptake of naked DNA (transformation). • Phages (transduction). • Plasmids (conjugation). • Integrative and transposable elements (transposition). • Cell fusion (in Archaea).
Transformation and competence • Many microorganisms can naturally take up DNA fragments from broken cells. • Inserted DNA can sometimes be recombined into the chromosome.
Phages - generalized transduction Many phages mistakenly package some host DNA. Resulting particle often cannot replicate, but does inject the foreign DNA. Homologous recombination can lead to integration of acquired DNA.
Phages - specialized transduction Some phages can lysogenize – integrate their DNA into the host chromosome. This integration is site-specific. This is often benficial to the host – protects from related phages and sometimes confers advantages (toxin genes in phages of C. diphteriae). The phage can later be induced to exit the chromosome and replicate (lytic cycle). Rarely the phage packages neighboring host genes, leaving some of its DNA behind. Thus, a phage can shuttle DNA between prokaryotes, or “contribute” phage genes to their genome.
Plasmids and conjugation • Plasmids are genetic elements that replicate independently of the host chromosome. • Exist as free (usually circular) DNA. • Generally do not encode essential genes. • Are spread among cells by cell to cell contact – conjugation, usually involving-plasmid encoded pili. • Host range varies from narrow to broad depending on replication machinery (and usually not the conjugation factors). • Some plasmids can integrate into the chromosome and subsequently their conjugation can mobilize parts of it. Integrated plasmids (episomes) can sometimes recombine with the host chromosome and exit with a few chromosomal genes.
Transposable elements (IS elemens and transposons) Genetic elements that can move within the genome. Contain a gene encoding a transposase flanked by inverted repeats and in the case of transposons also other genes. Usually integrate at specific sites. Are also found in phages and plasmids. Can mediate recombination and chromosome re-arrangement. = Inverted repeat
Transposable elements can jump from one microbe to another by either: • Being located on a mobile element such as phage or plasmid. • Having their own conjugation systems – Integrating Conjugative Elements (ICEs).
Integrons: Chromosomal elements that recruit gene cassettes by site-specific recombination
Clinical importance of LGT:Staphylococcus aureus as an example • Phage-encoded toxins: • Enterotoxin A - food poisoning. • Exfoliative toxin A - scalded skin syndrome. • Panton-Valentine leukocidin - severe skin infections and necrotizing pneumonia in children (mortality rate of 40%). • Plasmids - Antiseptic and antibiotic resistances. • Integron- Methicillin resistance. • Transposon-encoded vancomycin resistance (from plasmids of enterococci!). Based on Lindsay and Holden, 2004, (Trends Microbiol. 12:378-85)
LGT - driving adaptation and speciationFrom colitis to plague - genes acquired less than 20000 years ago “created” the species Yersinia pestis • Ymt – Toxin, also essential for flea colonization. • Plasminogen activator Pla, invasin essential for virulence by the subcutaneous route. • Following the new lifestyle – major gene loss and accelerated evolution formed a new bacterial species.
Factors influencing success of LGT • Evolutionary/Genetic distance between organisms. • Physical proximity of organisms. • Gene function: Frequently transferred: (Strong positive selection) • Resistance genes – Antibiotics, heavy metals, arsenic... • Virulence factors – Pathogenicity islands. • Metabolic pathways – Metabolic islands/plasmids. Rarely transferred: • Genes that are part of essential complexes, such as the translation machinery – “The complexity hypothesis”.
Acquisition of potentially useful genes • A totally new function. • A function already performed by a homolog – quasi duplication – sometimes followed by orthologous replacement. • A function already performed by a non-homologous gene –leading to either non-orthologous replacement or functional divergence.
Comparative genomics : There is much LGT but is it always adaptive ?
EVIDENCE FOR LGT: PROKARYOTIC GENOMES AS MOSAICS ---- --- “backbone” or “core” genes with common history “island” of genes with alien origin phage gene single alien genes Typical stretch of prokaryotic chromosome sequence
LGT and the Tree of Life conflicting views of its role in evolution Committed verticalists • LGT is rare and generally has little impact on evolutionary processes (Kurland). • A relatively stable core of genes is very rarely transferred, while most other genes can undergo LGT (Woese). • Since most if not all genes in a genome have been transferred at least once, a tree of life is meaningless (Doolittle). Enthusiastic lateralists
The Universal Tree of Life mitochondria chloroplasts LAST UNIVERSAL COMMON ANCESTOR (LUCA) Adapted from Doolittle, 1999 (Science, 284:2124-2128)
A core of genes showing the same true tree core genes LUCA (Last Universal Common Ancestor)
A web of life - no true tree and no LUCA (there are no species trees just gene trees) Adapted from Doolittle, 1999 (Science, 284:2124-2128)
How can we accurately assess the impact of LGT? . Lateral gene transfer and the nature of bacterial innovation. Ochman, Lawrence, Groisman. Nature. 2000;405(6784):299-304 Extent of LGT Differs between genomes
How can we detect LGT? • Phylogenetic trees. • Composition-based methods: G+C content, codon usage. • Phylogenetic discordance – atypical patterns of similarity to different organisms. • Distributional profiles.
Caulobacter Caulobacter Brucella Sinorhizobium Yersinia Yersinia Escherichia Brucella Escherichia Detecting LGT Tree based detectionComparing species tree to gene trees Sinorhizobium Salmonella Salmonella Gene Tree Species Tree(usually 16S)
Detecting LGT Trees are not always an option… • Making trees for all genes in a genome requires pipelines that are computationally intensive and still require much human intervention – Moore vs. Moore. • Inferring LGT by tree reconstruction requires at least four-five homologs.
Detecting LGT Tree-free methods • Composition-based methods: G+C content, codon usage... • Distributional profiles. • Phylogenetic discordance – atypical patterns of similarity to different organisms.
Fraction of LGT detected Phylogenetic depth of inferred event Phylogenetic depth of inferred event (antiquity) Compositional methods fail to detect many ancient events – ancient LGT is often underestimated G+C Content Codon usage (MM) Phylogenetic Discordance Distributional Profiles Ragan, Harlow and Beiko 2006, (Trends Microbiol. 14:4-8)
Which laterally acquired genes tend to be fixed in a microbial population? • Genes under strong positive selection. • Genes that form a functional cluster (Selfish operon theory…). • Genes that are not part of essential complexes (see Alon Wellner’s poster). • Genes with compatible codon usage to the new host (Medrano-Soto et al., 2004). • Acquiring a gene by LGT is up to six times more likely if an enzyme that catalyses a coupled metabolite flux is already present in the genome (Pal et al., 2005).
Acknowledgements W. Ford Doolittle Alon Wellner (see poster!) Mor Lurie Amitai Or Amir Kovacs Funding Research Networks Program in Bioinformatics (Israeli MOST/ Ministry of Foreign Affairs and the Ministry of National Education and Research of France.