Download
1 / 55
700 likes | 1.85k Views
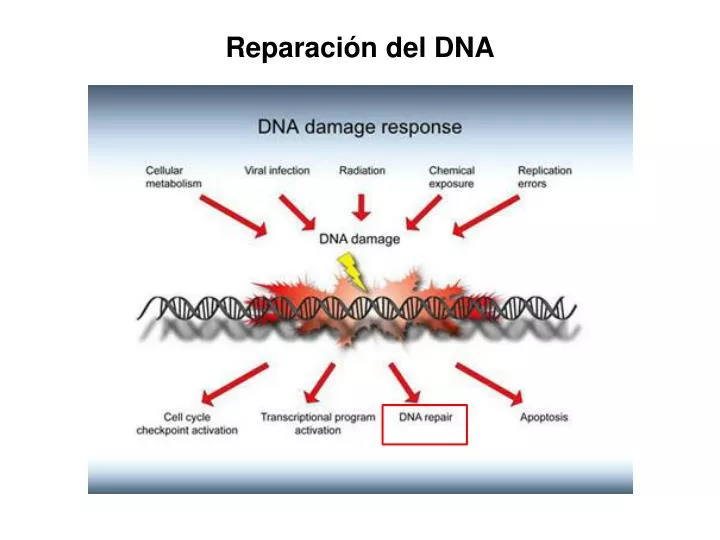
Reparación del DNA. Daño al DNA puede ocasionar mutaciones. Los daños en el DNA son minimizados por sistemas que los reconocen y corrigen. SISTEMA DE REPARACIÓN EXITOSO. SISTEMA DE REPARACIÓN X. Sin consecuencias. MUTACIÓN. Daño al DNA. Daño al DNA.

E N D
Daño al DNA puede ocasionar mutaciones • Los daños en el DNA son minimizados por sistemas que los reconocen y corrigen SISTEMA DE REPARACIÓN EXITOSO SISTEMA DE REPARACIÓN X Sin consecuencias MUTACIÓN Daño al DNA Daño al DNA
Daños espontáneos al DNA Sitios AP 5000-10000 al día inestable 100 al día
Daños inducidos al DNA La metil guanosina se aparea de forma incorrecta con la timina causando un cambio de G-C a T-A Radiaciones ionizantes: rayos X pueden causar rompimiento en el DNA
MECANISMOS DE REPARACION • 1- Sistemas de Reparación directos • Enzimas que revierten directamente el daño. • Por estos mecanismos se reparan: metilación de guanina, y en algunos vertebrados dímeros de pirimidína. No intervienen nucleasas ni ADN-polimerasas. • Fotoreactivación(ruptura de los dímeros de pirimidinas por acción de una fotoliasa (phr) activada mediante luz visible).
2- Sistemas de Reparación Indirecta • Hay intervención de nucleasas y ADN-polimerasas. Se necesita de la hebra “molde” perteneciente al mismo cromosoma o al homólogo. • Reparación por Escisión (BER , NER, MMR) • Reparación de nucleótidos (NER)aislados por lesión UV: necesita la otra hebra como templado (hasta 30 bp). Intervienen las endonucleasas uvrA,B,C y la helicasa uvrD. • Además de foto productos, repara lesiones voluminosas (bulky) que distorsionan la conformación del dúplex y que obstaculizarían la transcripción y replicación. • Reparación de bases modificadas (BER).- Repara casos de alteraciones puntuales en bases nitrogenadas (lesiones NO voluminosas) producidas por alquilación, oxidación o desaminación. Se origina un “sitio AP” y luego se retira el nucleótido “AP” y se re sintetiza la hebra)
Reparación por escisión de Bases (BER) 1) Iniciado por DNA glicosilasa específica reconoce el daño, corta la unión glicosílica entre base y azúcar y se forma el sitio AP 2) Sitio AP reconocido por AP endonucleasa (corte 5’ de AP). 3) Fosfodiesterasa (corte 3’). 4) DNA polimerasa rellena el gap DNApol I (E.coli), DNA pol (mamíferos). 5) DNA ligasa 2) Reparación Indrecta de Daño al DNA
Reparación por escisión de Nucleótidos (NER) Escinucleasa uvrABC realiza este tipo de reparación en dímeros de timina, otros fotoproductos y bases dañadas. Escinucleasa (246 kDa) está compuesta por tres subunidades (A, B y C) UvrA (dímero con actividad de ATPasa)se une al DNA en la región dañada. UvrB/UvrC tienen actividad de endonucleasa y corta en los lados adyacentes de la cadena liberando un oligonucleótido La región “vacía” es rellenada por una DNA polimerasa I y sellada por una DNA ligasa. UvrD E.coliSistema Uvr ABC: Remoción de 12nt EucariontesRemoción de 24-29 nt
Reparación post- replicativa • Reparación del apareamiento (MMR) (“mismatch repair”): • Su principal tarea es remover bases mal aparadas y pequeños “loops” introducidos por inserciones / deleciones durante la replicación • Reduce los errores de replicación de 10-7 a 10-10 pb / replicación
Reparación de bases mal apareadas(MMR) E.coli genes mut S, L, H Reemplaza hasta 1kb Metilación diferencial (dam, dcm) MutS reconoce el mismatch MutH distingue ambas cadenas Corte en GATC en la cadena no metilada Mut L coordina actividad de Mut S y H En eucariotas homólogos de proteínas Mut
… Reparación post- replicativa • Recombinación Homóloga (HR) • Reparación de ambas cadenas • Usa ADN homólogo como templado y es altamente exacto • Más activo durante la Fase S y G2 • Unión de extremos no homólogos (NHEJ) • Reparación de ambas cadenas • No usa ADN templado y generalmente se pierden algunos nucleótidos. • Más activo en la Fase G1
polIV Respuesta SOS polII Pol V
Mecanismos de reparación en Eucariontes (NER) Participan > de 25 proteínas Reconoce el daño 3 síndromes XP-B XP-D Helicasas que forman parte de TFIIH Abre las cadenas XP-A confirma la presencia del daño XP-G; XP-F endonucleasas Corrige el error
XPA-XPG CSA y CSB
RECOMBINACIÓN Recombinación homóloga Recombinación no homóloga
Recombinación homóloga Recombinación entre cromátidas hermanas Recombinación entre cromosomas homólogos
El modelo Holliday (c) strand exchange takes place between the chromatids (d) ligation occurs yielding two completely intact DNA molecules (a) pair of chromatids (b) a single strand cut is made in each chromatid
Resolución de las uniones Holliday Holliday junction
El modelo de la cadenadoble fragmentada
Secuencia Chi • Estimula la recombinación en forma direccional • 5´-GCTGGTGG-3´ • Octámero sobre-representado en el genoma de E. coli. Existen • aprox 1,008 sitios Chi. • Aparecen en promedio cada 4.5 kpb. • 75% de los sitios Chi están orientados hacia el origen de la replicación
RecBCD 134 kDa 129 kDa 67 kDa
Translocación bipolar RecD RecB = velocidad ≠ velocidad
3´ Vía RecBCD 5´ Ocasionalmente + frecuente
Proteína RecA 37kDa Aprox 8,000 a 10,000 Aumenta a 70,000 en SOS 3 nucleótidos/monómero que se extiende en dirección 5´--3´
Enzimas bacterianas que catalizan la recombinación RecA Intercambio de cadenas de DNA RuvA, B Migración de los entrecruces. RuvA es tetramérica y tiene alta afinidad por HJ independiente de la secuencia. RuvB es hexamérica y tiene afinidad por DNA. Tiene actividad de ATPasa que es estimulada por unión a RuvA. RuvC Nucleasa resuelve entrecruzamientos