Download

1 / 25
250 likes | 270 Views
This text reveals the elucidation of electron carriers, inhibitors, and ATP synthesis coupling in mitochondria. Chemiosmotic model's paradigm shift, proposed by Peter Mitchell, explains electron flow and ATP synthesis coupling in bioenergetics.

E N D
The order of the electron carriers have been elucidated by using inhibitors and artificial electron acceptoers, as well as measuring the standard reduction potentials and oxidation kinetics
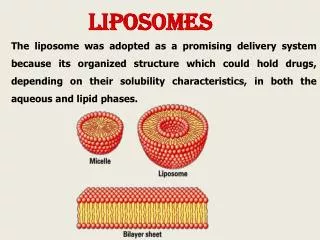
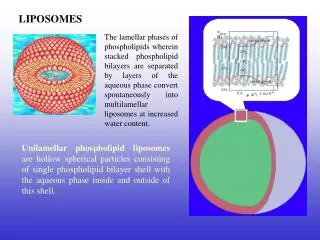
Functional complexeshave been successfully separated via ion-exchange chromatography, with the activity and components of each being characterized in detail. Purified complexes can be studied by reconstituting into liposomes. Supercomplexes have been revealed
Electron transfer to O2 was found to be obligatorily coupled to ATP synthesis (from ADP + Pi) in isolated mitochondria. Respiration is tightly coupled to the synthesis of ATP!
ATP synthesis does not occur in isolated mitochondria suspensions when an electron donor is missing or electron transfer is blocked. The e- transfer measured by O2 consumption. ATP synthesis
Electron transfer could not be detected in isolated mitochondria suspensions when ATP synthesis does not occur, unless certain weak hydrophobic acids were added. Electron transfer and ATP synthesis are obligatorily coupled: neither occurs without the other.
The coupleing between electron transfer and ATP synthesis was found to occur via a proton gradient across the inner membrane of mitochondria
It was widely believed that ATP synthesis occurs by chemical coupling (1940s-1960s) • High energy intermediates similar to 1,3-bisphophoglycerate was once proposed to be produced first from the electron flows on both the mitochondrial and chloroplast membranes. • Phophorylated protein intermediates (as formed in the action of phosphoglycerate mutase and phosphoglucomutase) were also hypothesized. • But neither were ever revealed despite intense efforts by many investigators over several decades.
BUT:It is difficult to explain the remarkable uncoupling action of 2,4-dinitrophenate, other chemically unrelated reagents, and the physical membrane-lytic treatments
The chemiosmotic model was proposed by Peter Mitchell in 1961 to explain the coupling of electron flow and ATP synthesis(The term Chemiosmotic emphasizes that chemical reactions can be coupled to osmotic gradients.) Mitchell, P. 1961, Nature, 191:144-148. A “paradigm” shift in understanding bioenergetics!
The chemiosmotic theory of Mitchell:e- flow and ATP synthesis are separate events, coupled via a transmembrane H+ gradient!
The chemiosmotic model was strongly supported by experimental observations • First, a closed membrane system is essential for ATP synthesis but not for the electron flow (tested with detergent or physical shearing).
Second, Some hydrophobic weak acids (e.g., DNP) were found to be able to uncouple ATP synthesis from electron flow. Electron transferring continued Even when ATP synthesis is stopped.
Hydrophobic weak acids are able to carry protons across the membraneThe proton gradient is thus dissipated by such “uncouplers” DNP was once used as a “diet pill” to induce weight loss.
Third, artificially imposed protongradient and charge imbalance across the inner membrane of mitochondria alone(in the absence of an oxidizablesubstrate) was found to be able to drive ATP synthesis! pH 9 pH 7 PH 7
The chemiosmotic theory unified the apparently disparate energy transduction processes as oxidative phosphorylation, photophosphorylation, active transport across membrane and the motion of bacterial flagella.
Abundant knob-like protruding structures were observed on the matrix side of the inner membrane of mitochondria. ATP synthesis did not occur when the “knob” (F1) was removed and resumed when added back (reconstitution). Electron micrograph of the inner membrane of mitochondria (Racker, 1960)
Similar “knobs” were also observed on other membranes by EM. The thylakoid membranes The inside-out vesicles from E. coli
Unexpected results from in vitro studies Of F1 ATPase! The18O exchange experiment: the Gˊ0 for ATP synthesis on purified F1 is close to zero!(Paul Boyer,1970s )ATP formation/hydrolysis are readily reversible in the active site of purified F1 (in the absence of a proton-motive force!) “uncouplers” exhibit no effect on such exchange when assayed with isolated mitochondria! Boyer (in 2000)
The proton-motive force was proposed to be needed for releasing ATP from ATP synthase, not needed for the formation of ATP (Boyer et al., 1973, PNAS, 70:2837-9. Measurement of Kd values: ATP has a much higher affinity than ADP to the enzyme (10 -12 M vs 10–5 M).
Stationary unit (stator) F1: ATP synthase, “a splendid molecular machine”, comprises a proton channel (Fo) and an ATPase (F1) Catalytic Proton-conducting The rotor: the c ring and the stalk; the stator: the remainder. Fo: ab2c10-14
The binding-change model was proposed by Paul Boyer to explain the action mechanism of ATP synthaseDownhill proton movement through Fo is proposed to drive the rotation of the c-subunit ring and the asymmetrical subunit, which will cause each of the three subunits to interconvert between three conformations.