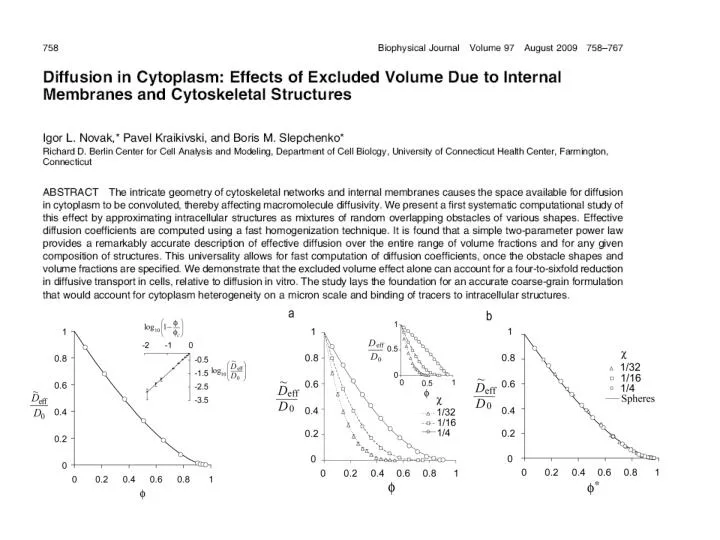
Download
1 / 7
70 likes | 199 Views
Quantifying a pathway: Kinetic analysis of actin dendritic nucleation Pavel Kraikivski and Boris M. Slepchenko. Progress in uncovering reaction networks that underlie important cell functions lays groundwork for quantitative identification of protein interactions involved in the pathways.

E N D
Quantifying a pathway: Kinetic analysis of actin dendritic nucleation Pavel Kraikivski and Boris M. Slepchenko Progress in uncovering reaction networks that underlie important cell functions lays groundwork for quantitative identification of protein interactions involved in the pathways. Direct experimental determination of rate constants is not always feasible, particularly in vivo. Reaction parameters are often inferred from multiple pieces of information about dynamics of the system as a whole, using kinetic analyses based on appropriate mathematical models. The issue critical for success of this approach is whether the available experimental data are sufficient for unique parameterization of the network and if not, what additional experimental measurements are required. We analyze the problem of quantifying a reaction pathway in the context of actin dendritic nucleation, an Arp2/3-mediated mechanism that enables rapid changes of cell shape in response to external cues.
Experimental data Beltzner & Pollard (2008) Mullins et al. (1998) Polymerization of 5.3 mM pyrene-labeled actin accelerated by Arp2/3 complex at following concentrations (right to left): 0 mM, 0.037 mM, 0.15 mM, 0.58 mM, and 2.3 mM. Solid lines are kinetic simulations. The concentration of polymerized actin was plotted versus time for pyrene actin assembly data (solid lines) and simulations performed using the program Virtual Cell (dashed lines). Reactions contained 4mM actin, 1mM VCA and the following concentrations of Arp2/3 complex: black, 0 nM; red, 2 nM; orange, 5nM; light green, 10nM; dark green, 20nM; blue, 50nM; and purple, 100 nM. Pantaloni et al. (2000) Noisy lines are pyrene-fluorescence measurements, dotted lines are calculated time courses. Reactions contained 2.5 mM G-actin, 0.6 mM WA and the Arp2/3 complex at the following concentrations (right to left): 0, 1.76, 3.52, 7.2, 14.3, 28.5, 56, 71 nM
kon, koff kact kbind Mathematical model Model Arp2/3-mediated actin nucleation pathway
a. Nucleation is limited by step 1: b. Nucleation is limited by step 2: Fitting data by Beltzner & Pollard indeed yields Experiment (Beltzner & Pollard, 2008): Data cannot be fitted on the assumption that step 1 is rate-limiting. Available data are insufficient for unambiguous pathway parameterization Idea: are the data consistent with the presence of a rate-limiting step? Fitting data by solving full model numerically with nucleation rates limited by step 2 (blue lines) or by step 3 (red lines). (a) Beltzner & Pollard, 2008; (b) Mullins et al., 1998; (c) Pantaloni et al., 2000.
Results of Pantaloni et al.(2000) are consistent with the side branching mechanism Experiment: data by Pantaloni et al., Nat Cell Biol (2000) Results from simulations based on the side-branching mechanism Expected length dependence is obscured by high number density of the seed filaments used in the experiments (tens of pM). Under these conditions, the initial dynamics of F-actin (when the process is not yet significantly influenced by branching) are dominated by polymerization at the barbed ends and therefore differ only slightly for varying seed lengths. As a result, there are similar amounts of F-actin when the Arp2/3-mediated nucleation comes into effect, thus producing similar dynamics overall.
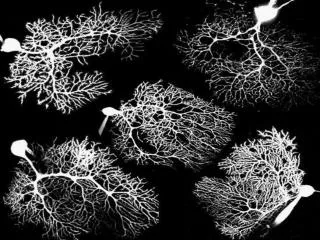
How to distinguish between the two possible rate limiting steps? We propose to take advantage of the fact that the nucleation limited by binding of the nucleating complex to a filament occurs faster for larger G0, whereas the rate of nucleation limited by activation (step 3) is largely independent of the initial concentration of monomers G0. This yields qualitatively different dependences of the average F-actin density on the initial monomer concentration: an increasing one, for the binding-limited nucleation, and a decreasing one, if the nucleation is limited by activation. The F-actin density could be measured using an experimental setup described in Haviv et al., PNAS (2006), in which aster-like structures of actin meshwork were observed. Spatial stochastic simulations of actin polymerization yield qualitatively different dependences of F-actin density on initial G-actin concentration for two rate-limiting steps: (a) binding of the nucleation complex to a filament; (b) activation of the bound complex. The plots of average F-actin density reflect values obtained from the snapshots using ImageJ software. Aster-like structure of polymerized actin observed by Haviv et al. PNAS (2006).