Download

1 / 40
430 likes | 934 Views
Part 1: pH, Buffers & Amino Acids. Introduction Cells made up of many organic cpds: nucleic acids polysacharrides - chains of sugars lipids - chains of fatty acids proteins => contain weak acidic & basic groups => change in pH --> change in ionisation state of the groups

E N D
Part 1: pH, Buffers & Amino Acids Introduction Cells made up of many organic cpds: • nucleic acids • polysacharrides - chains of sugars • lipids - chains of fatty acids • proteins => contain weak acidic & basic groups => change in pH --> change in ionisation state of the groups Solution? Use buffers to resist changes in pH (pg.1)
Objectives • What is pH? • What is strong/weak acid & base? • What are buffers? • What are amino acids?
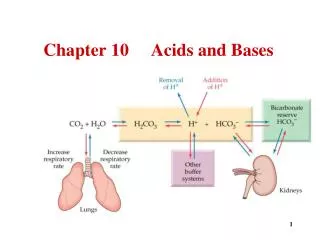
1. pH, Acids & Bases Introduction (pg.2) • Bronsted & Lowry: Acid is a molecule that can ionise to release a hydrogen ion: HA <--> H+ + A- • In aqueous solution, hydrogen ion exists as hydronium ion • HA + H2O <--> H3O+ + A- (conjugate acid) (conjugate base)
1.1 What is pH & Kw? Definition of pH (pg2) • Negative logarithm of the concentration of free hydrogen/hydronium (H+ / H3O+) ions in a solution pH = -log10[H+ ] • If [H+] = 10-3M => log10 [H+ ] = -3 => pH = -log10 [H+ ] = 3 Ionisation of Water, Kw (pg3) H2O + H2O <--> H3O+ + OH- • Dissociation constant of a solution Kw = [H3O+] [OH-] = 1 x 10-14 M • For Water: [H3O+] = [OH-] = 1 x 10-7 M => neutral pH • Table 1: the pH scale (pg4)
1.3 Acid Dissociation Constant, Ka • A measure of the extent an acid dissociates in aqueous solution (pg3) HA + H2O --> H3O+ + A- Ka = [H3O+] [A-] [HA] => greater dissociation => larger Ka => • pKa = -log10 Ka
1.4 pH and pKa • Henderson-Hasselbalch Equation (pg4): pH = pKa + log10([base]/[acid]) = pKa + log10([A-]/[HA]) (i) RCOOH <=> H+ + RCOO- ---> pH = (ii) RNH3+ <=> H+ + RNH2 --> pH = • For weak acid, pH = pKa when [base] = [acid] => acid is 50% dissociated
2.1 Strong Acid & Base HA + H2O --> H3O+ + A- (pg4) • Completely dissociates in water => [H3O+ ] = [HA] => pH = -log10 [H3O+] = -log10 [HA] (i) pH of 0.1M HCl = (ii) pH of 5 x 10-2 M H+ = (iii) pH of 0.01M NaOH = (iv) pH of 0.030M OH- =
2.2 Weak Acid & Base • Ionisation not complete (50%) (pg5) • pH (of weak acid) = 1/2(pKa - log10[HA]) • pH (of weak base) = 7 + 1/2(pKa - log10[base]) pH of 0.1M acetic acid (Ka = 1.73 x 10-5 M) = pH of 0.1M NH4+ (Ka = 1.8 x 10-5 M) =
3. Buffers • Weak acid-base pairs (pg6) • To maintain pH of a solution • Biological rxns --> acidic or basic products => inhibits enz rxn (pg5) • Buffers ‘mops’ up free H+ or OH- ionic products • Buffers cannot withstand large amounts of acid or alkalis • Effective buffer is one that can buffer between pKa + 1 and pKa - 1 (pg6) eg. acetic acid-acetate buffer at pH phosphoric acid-phospate at pH ammonia-ammonium at pH
3.1 Acetic acid-Acetate Buffer • Acetic acid solution has mixture of undissociated acetic acid, acetate ions & hydronium ions (pg6) CH3COOH + H2O <=> CH3COO- + H3O+ + OH- --> 2H2O • Each OH- ion neutralise one H3O+ ion • Acetic acid ionise to produce more H3O+ ions to neutralise OH- => pH of solution remains relatively constant: • End point is when there is no more acetic acid --> pH increase • The reverse happens when when a small quantity of H+ is added
4. Amino Acids • Free amino acids consist of (pg9) • an a-amino group(NH2) • an a-carboxyl group (COOH) • a side chain group (R) • The NH2 & COOH group (& sometimes the R group) can gain or lose protons - depend’g on the concentration of protons (pH) of the solution => AA can be used
4.1 Ionisable forms of Amino Acids • AA with non ionisable R group can exist as (pg9, 10) • cation (overall +ve charge) - at low pH • anion (overall -ve charge) - at high pH • zwitterion (+ve = -ve charge) - at isoionic/isoelectric point • AA with ionisable R group can exist as (pg10) • cation • zwitterion • intermediate ion (overall +ve or -ve depend’g on R group) • anion
4.2 Fractional charge of ionisable group • Tells the percent of the group that carries a charge at any one time for a given pH (pg12) eg. (i) FC of -0.5 for carboxyl group for a given pH => 50% of the carboxyl grps carries a -1 charge & 50% carry no charge (ii) FC of +0.3 for an amino group for a given pH => 30% carry +1 charge (RNH3+) & 70% carry no charge (iii) If at a given pH an AA has 50% carboxyl grps charged -1 and 30% charged +1 => overall charge =
cont. Fractional charge • Can calculate FC on any ionisable group in an AA for any pH if you know the pKa (pg12) FC = charged species x magnitude & sign (+/-) total species • Tables (a) & (b) pg 12 & 13
4.3 Free AAs & ‘Bound’ AAs • pKas for free AA side chains differ by up to + 1 pH from pKas of Aas in a protein (pg14) • Acid-base properties of an AA residue in a polypeptide/protein depends upon • neighbour’g residues can influence the properties of a side chain • a residue buried within the interior of a protein molecule and the same type of residue located on the surface of the protein may have a different pKa
4.4 Effects from changes in pH • pH determines the types of ionic forms & their proportions in an environment (pg14) • The different ionic forms determine other physical & chemical properties of AAs eg. optical rotation, uv absorbance, metal chelating activity, • These properties are also affected by changes => pH influences the charge of amino acids & hence also biomolecules --> thus affect’g its function
Part 2: Proteins Introduction • Proteins are components of all living things • Contain the elements C, H, O, N & S • Polymer of amino acids linked by peptide bonds • Polypeptides of more than 50 AA • Simpleprotein - made up entirely of AA residues or Conjugated protein - has another non-AA component (prosthetic grp) incorporated • Fibrous protein eg. collagen or Globular protein eg.
1 Molecular Properties of Proteins • Protein Structure (pg55) • 1o structure - long, continuous, • 2o structure - linear chain folds & turns on itself • 3o structure - chain aggregates to form a 3-D structure: • 4o structure - aggregation of 1 or more chains • Folding: Aas with hydrophilic R grps on the outside & Aas with hydrophobic R grps are inside the molecule • Shape & size - covalent peptide, disulfide bonds; hydrogen bonds, • Changes in physical, chemical & bioogical properties mainly due to changes in non covalent bonds • Prots have different types & proportions of Aas => differ in shape, size, charge, solubility, stability
cont. Molecular Properties • Protein denaturation (pg56) • reversible or irreversible change in 2o, 3o, 4o • by a chemical eg. SDS, urea, high acid/alkaline • by physical process eg. rapid stirring, heat • Absorption of light by Proteins • absorbs uv light at 250nm to 300nm • measure protein concn • Chemical assays of Proteins by spectrophotometry • Biuret method (540nm) • Lowry method (750nm) • Coomassie Brilliant Blue or Bromocrsol Green
2. Effect of pH on Structure & Function • Aas bound to each other by peptide bonds - a-amino group of 1 AA linked to the a-carboxyl group of another AA (pg57) • Ionisable groups: • the a-amino & the a-carboxyl grps at the ends of the chain • the side grps • Changes in pH (pg58): • proteins can act as buffers • can alter proportion & distribution of +ve & -ve charges => may cause changes in shape, orientation => may affect structure, stability, function
2.1 Isoelectric Point (pI) • the pH at which the protein has no net charge • acidic protein: low pI & negatively charged • basic protein: high pI & positively charged • protein in a solution wh pH > pI => will bear a net -ve • protein in a solution wh pH < pI => will bear a net +ve
2.2 Effect of pH & Ionic Strength on Protein Solubility • Adjust pH or ionic strength -> alter distribution of +ve & -ve charges on the molecule -> affect protein’s solubiliy (affinity with water) (pg59) • Isoelectric point of a protein => pH where protein is least soluble (pg59; Fig2, pg60) • Ionic strength - measure of the total charge of ions in solution • increase in IS of solution • low salt concn => increase solubility of protein but • very high salt concns => solubility decreases (see pg 59)
3. Separation Methods Chromatography (pg60) • separates a mixture into its components or isolates one component fr a complex mixture • Stationary phase (SP) - solid, liquid or solid/liqd mixture - immobilised • Mobile phase (MP) - liquid or gas wh flows over or through the SP • Elution: movement of the MP over or through the SP • Components of a mixture distribute themselves bewtn the MP • Kd (distribution coeffcnt) = concn of component concn of component in MP
cont. Chromatography Classification based on method used in separation • Physical system - column • Phase - liquid or gas • Chemistry - adsorption, filtration, ion exchange, affinity • Single step (batch) • Multi-step (column)
3.1 Ion Exchange Chromatography • separates molecules based on their net charge (pgs62, 63) • stationary phase: -vely or +vely charged functional groups covalently bound to a water-insoluble • Cation (acidic) exchanger: SP -ve functnl grp, binds mobile +ve ions • Anion (basic) exchanger: SP +ve functnl grp binds mobile -ve ions Matrix--C- :: A+ + X+ <--> Matrix--C- :: X+ + A+ ion exchange resin counterion sample ion • Molecules of opposite charge interacts with the SP; neutral ions & ions of same charge are eluted • Molecules with large charge will interact strongly; small charge interact moderately
3.2 Gel Filtration Chromatography • Gel permeation or Size exclusion chromatography (pgs 64, 65) • Separates molecules based on molecular size • SP: polymers of organic cpds cross-linked to create a 3-D porous matrix • Size of pores in gel determined by degree of cross-linking • Large molecules cannot enter (excluded) the gel matrix -> first to elute • Small molecules permeate pores -> last to elute • Elute molecules in decreasing size
3.3 Electrophoresis • Proteins are charged at a pH other than their pI => exist with net -ve or +ve charge in solution (pg68) • Proteins move thru’ a solution under an electric field: cations travel toward cathode anions travel toward anode => Separate proteins based on electrophoretic ability • Solid matrix to support molecules during separation => GE
cont. Electrophoresis • Buffers used (pg69): • Non-dissociating buffer separates proteins by charge, molecular wt & size - prots retain 3-D struc • Dissociating buffer (SDS) separates proteins by molr wt & size (not charge) => can estimate molr wt of unknown when compared with prots of known mol wt • Continuous system uses the same buffer ions in sample, gel & electrophoresis • Discontinuous buffer system uses difft buffer ions for the gel & electrophoresis
4. Protein Purification • Preparative Purification Techniques (physical & chemical) (pg70) • isolate proteins in relatively large amounts • purify protein • evaluate purity (using analytical techns) at each step eg. enzyme activity • estimate total amount of protein by standard protein assay
cont. Protein Purification • Measurement of Enzyme Activity (pg71) A + B --> C + D => C absorbs light at nm but not Ab, B & D => measure production of C at nm • a buffered rxn mixt contain’g substrate(s) & a known volume • measure (spectrophotometrically) the production of a product • calculate the rate of change • enzyme activity in International Units (I.U.) - amt of enzyme wh catalyses conversion of 1umol substrate/min under defined rxn conditions
cont. Protein Purification • Specific Activity (SA) (pg72) • by assay’g for enzymic activity & absolute prot • SA (IUmg-1) = enz activity per ml sample(IUml-1) total prot concn (mgml-1) • Purification Factor • proportion of the total prot wh is made up of the prot of interest • Purification = specific activity of sample (IU per mg) specific actvty of start’g material (IU per mg) • Yield = Total units of enz actvty in sample x 100 Total units of enz actvty in start’g material
cont. Protein Purification • Factors Causing Loss of Enzyme Activity (pg73) • Physical loss (eg. not all precipitated) • Inactivation or denaturation • frothing of solution by vigorous agitation • inappropriate pH, temp. • heavy metal ions wh inhibit • breakdown of enz • Analysis of purity (pg74) • indirectly, by determining specific activity • directly by separating prots
Part 3. Enzyme Activity & Kinetics • Enzyme assays • estimate amount of active enz present in cell • monitor purification of enzymes • provide info on catalytic mechanisms & physiological role of enz • Enzymes (pg98, 99) • catalysts to a rxn => affect only rxn rate • 3-D struc controlled by many factors: pH, salts, • changes in temp - alter rxn rates, activity, struc • salts cause denaturation (maybe reversible) • heavy metals alter struc
cont. Enzymes • E + S == ES <==> EP == E + P (pg98) v1 = k1[E][S] formtn of enz-substr v2 = k2[ES] reformtn of free enz & substr v3 = k3[ES] formtn of product v4 = k4[E][P] reformtn of enz-prodt complex In steady state equilibrium, v1-v2 = v3-v4 & if all product is removed/does not recombine with enz => k1[E][S] - k2[ES] = k3[ES] => (k2 - k3)/k1 = [E][S]/[ES] where (k2 - k3)/k1 is Km (rate constant or Michaelis constant = measure of enz actvty) • Enzs fr difft sources (but same function) may have difft Km
1. Active Sites • The particular part of the enzyme structure which specifically binds to a substrate (pg99) • Enzyme does not react => it brings substrate into proper alignment/configuration for spontaneous rxn or rxn with another substance • Rxn proceeds by random kinetic action of molecules bumping into eah other => enzyme align substrate to facilitate rxn • When enzyme is in ideal configuration -> rxn proceed -> overall rate of activity dependent on substrate concn. • Maximum rxn rates at controlled conditions: optimal pH, salt envmnt, temp, presence of cofactors or co-enzymes, sufficient substrate to saturate all enzyme
1.1 Michaelis-Menten Plots • Plot enz activity /velocity of rxn (y-axis) vs substrate concn • Velocity of rxn = Initial velocity measured at each substrate concn. • From plot, as concn of substrate increases, velocity increases & approaches the maximum rate • At Vm, all enzyme molecules are complexed with substrate => any additional substrate has no effect on rate • Value of Vm dependent on enzyme concn & substrate concn
1.2 Michaelis-Menten Equation • Michaelis-Menten equation: v = Vm[S] (pg102) Km + [S] • Km (Michaelis constant) = rate constant = concn of substrate wh will give exactly 1/2 Vm when reacted with an enz with optimum pH, temp • However, difficult to obtain Vm accurately fr M-M plots => use Lineweaver/Burke
1.3 Lineweaver/Burke equation 1/v = Km + [S] = Km + [S] Vm [S] Vm [S] Vm [S] 1/v = Km + 1 Vm [S] Vm => straight line eqn • y = 1/v; x = 1/[S]; slope (m) = Km/Vm; intercept (b) = 1/Vm • easily obtain Km & Vm
2. Specific Activity • Enzyme units per mg enzyme protein (pg103) • An enzyme unit catalyse transformation of 1umole substrate per minute under specific rxn conditions (pH, temp & substrate concn) • Specific activity (SA) relates the enz units to the amt of prot • To obtain SA, measure amount of prot & kinetically measure
3. Enzyme Inhibition • If a molecule interferes with the binding of enz to substrate => inhibit the activity of the enz (pg104) • Competitive Inhibition: inhibitor molecule binds to same active site => No change in Vm, Km changes (req more substrate to compete) • Non-competitive Inhibition: Inhibitor binds to another site on enz & alters struc of enz or blocks access => Vm change (enz removed fr rxn) but Km no change • Uncompetitive Inhibition: has effects on both active site & allosteric site => Vm & Km change