Download
1 / 86
880 likes | 1.16k Views
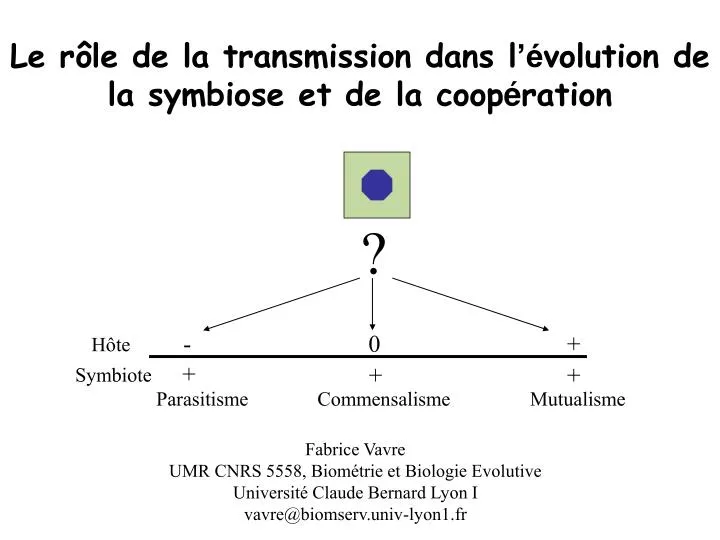
?. -. 0. +. H ô te. +. +. +. Symbiote. Parasitisme. Commensalisme. Mutualisme. Le r ôle de la transmission dans l ’é volution de la symbiose et de la coop é ration. Fabrice Vavre UMR CNRS 5558, Biométrie et Biologie Evolutive Université Claude Bernard Lyon I

E N D
? - 0 + Hôte + + + Symbiote Parasitisme Commensalisme Mutualisme Le rôle de la transmission dans l’évolution de la symbiose et de la coopération Fabrice Vavre UMR CNRS 5558, Biométrie et Biologie Evolutive Université Claude Bernard Lyon I vavre@biomserv.univ-lyon1.fr
Coopération Une large définition de la symbiose • Interaction durable entre deux espèces différentes (De Bary, 1879) Qu’entend-on par symbiose ? Toft (1991)
Environnement interactions GGE Génotypes des partenaires • Une association peut se déplacer le long de ce continuum Le continuum parasitisme-mutualisme • Parasites et mutualistes partagent de nombreuses caractéristiques - 0 + Hôte + + + Symbiote Parasitisme Commensalisme Mutualisme
Quelques questions qui émergent • Comment les interactions évoluent le long de ce continuum et quelles forces évolutives importent ? • Une question reliée: quelle est la stabilité du phénotype étendu, et quels facteurs sont importants ? • L’évolution de la coopération: un paradoxe ? • Qu’est ce qui rend l’évolution de la coopération possible ? • Pourquoi le mode de transmission est important ? • La réponse est-elle aussi simple ?
Host Pespective Cost: c Benefit: b Bh= f(Gh, Gs, E) and Ch=f(Gh, Gs, E) Une perspective dynamique de la symbiose • La sélection agit pour maximiser le bénéfice et pour minimiser le coût
Une perspective dynamique de la symbiose Symbiont Perspective Cost: c Benefit: b Bs= f’(Gh, Gs, E) and Cs=f’(Gh, Gs, E) • La sélection agit pour maximiser le bénéfice et pour minimiser le coût
Le paradoxe de la coopération • Mutualisme lorsque Bh>Ch ET Bs>Cs • Comment combiner des pressions sélectives a priori antagonistes? • Pourquoi les partenaires continuent à coopérer, alors qu’ils pourraient tirer un avantage à prendre le bénéfice sans payer le coût (évolution de la tricherie) ?
Le paradoxe de la coopération • « Natural selection cannot possibly produce any modification in any one species exclusively for the good of another species » C. Darwin • « The modern version of Darwin’s criterion is that the genes underlying a cooperative trait must themselves benefit disproportionately if they are to increase in frequency. » J.L. Sachs et al.
Gènes partagés = kin selection • Byproduct benefits • Directed Reciprocation • Partner fidelity feedback • Partner choice
Intraspécifique uniquement: la kin selection • La règle de Hamilton: rB>C Coût pour l’individu focal apparentement Bénéfice pour le receveur Un coût sur la fitness directe (altruisme vrai), mais une augmentation de la fitness inclusive
By-product mutualism Pas de coûts, seulement des bénéfices ! Syntrophisme L’origine de nombreux systèmes coopératifs ?
Mais ceci ne s’applique pas tout à fait à ce qui nous intéresse : L’altruisme réciproque
Le dilemme du prisonnier et la sélection de la coopération sans apparentement • Une seule interaction: sélection de la défection • Interaction répétées et « mémoire » : Défection et « Tit for tat » sont des ESS • La coopération peut être sélectionnée lorsqu les coopérateurs se rencontrent plus fréquemment qu’attendu Axelrod & Hamilton, 1981
“Partner Fidelity Feedback” :Le couplage des fitnessl’alignement des intérêts Bx augmente quand By augmente Un avantage direct à être plus coopératif Pas besoin de mémoire !
Verticale (de parent à descendants) G0 G1 Transmission et fidélité au partenaire Horizontale (contagieuse)
Transmission mixte et évolution expérimentale Une association : transmission controllée Seulement verticale Seulement horizontale G0 G1 Reduction de la virulence Sélection de la coopération Augmentation de la virulence Selection du parasitisme
Selection of benevolence in a host parasite system.Bull et al. 1991 • Le système : E. coli et un phage filamenteux (f1) • Phages non lytique, mais diminue la croissance cellulaire • Deux régimes de sélection : vertical, horizontal Des résultats retrouvés sur différentes associations hôte-parasite
A shift to parasitism in the jellyfish symbiont Symbiodinium microadriaticum.Sachs & Wilcox, 2006 • L’hôte : la méduse Cassiopea xamachana procure de l’azote et des nutriments inorganiques • Le symbiote S. microadriaticum procure des photosynthétats
A shift to parasitism in the jellyfish symbiont Symbiodinium microadriaticum.Sachs & Wilcox, 2006 TV TH
A shift to parasitism in the jellyfish symbiont Symbiodinium microadriaticum.Sachs & Wilcox, 2006 - TH sélectionne des symbiotes plus virulents, mais qui produisent plus de stades dispersants
G0 G1 Transmission Verticale et fidélité au partenaire • La corrélation de fitness entre l’hôte et le symbiote est inhérente au système • Une limitation dans les bénéfices apportés au partenaire affecte directement la fitness • Très resistant à la tricherie • Quelques exemples: mitochondries et plastes, symbiotes d’insectes • Une stabilité qui peut durer des millions d’années !
+ (500+200)/2-200=150 500-200=300 200-100=100 Et le gagnant est : (500+200)/2-100=250 Un facteur supplémentaire : le mixage des génotypes symbiotiques
Goulôt d’étranglement durant la TV uniparentale Mixage des génotypes lors de la TH Infections multiples, niveau de sélection et mode de transmission • La sélection intra-hôte favorise l’évolution de la tricherie • L’intensité de la sélection dépend de l’apparentement entre symbiotes au sein de l’hôte • Limitation de la competition intra-hôte par la TV conduit à une plus forte stabilité
Résumé • TV aligne les interêts des partenaires en couplant leurs fitness (partner fidelity feedback) • TV limite les infections multiples et la competition intra-hôte • Les symbioses à TV sont plus stables que les symbioses à TH • Pourquoi observe-t-on autant de symbioses mutualistes à TH ?
transmission Horizontale et mutualisme • Population visqueuses • Fidelité au partenaire augmente • Limite la compétition intra-hôte • Choix du partenaire Les génotypes les plus coopératifs sont sélectionnés Pas besoin d’interactions répétées
transmission horizontale • Mixage des partenaires : très sensible aux tricheurs • Mécanismes de renforcement de la corrélation de fitness entre partenaires • Choix du partenaire • Pourquoi la TV n’a pas été sélectionnée dans ces systèmes ? • Impossible? • L’expression de l’association dépend des conditions environnementales
« Conditional outcomes » et TV Infecté by H. defensa Non-infecté Oliver et al., 2005
« Conditional outcomes » et TV Oliver et al., 2008 • L’avantage dépend de la densité en parasitoïde • La TV ne permet pas d’ajuster la symbiose à des conditions fluctuantes
Est-ce aussi simple ?Transmission Verticale et reproduction sexuée père mère G0 • Les mâles sont des voies sans issue (rompt la fidelité au partenaire) • alignement des interêts seulement partiel avec le génome nucléaire (il reste de la TH au niveau du gène) G1
La sélection sur les symbiotes transmis maternellement L’infection se répand lorsque : Wi x SRi > Wn x SRn
La stérilité mâle cytoplasmique induite par les mitochondries • Gynodioécie: co-existence d’hermaphrodites et de plantes mâle-steriles • Stérilité mâle induite par les mitochondries • Les ressources qui servent normalement à la formation des gamètes mâles sont ré-allouées à la production de gamètes femelles (qui transmettent les mitochondries !) • Conflit entre les compartiments nucléaires et cytoplasmiques
G1 Les conflits Nucléo-cytoplamsiques G2 Conflits très forts qui sélectionnent pour des gènes de résistance nucléaires - Evolution de la restauration - Limiter la transmission des symbiotes (uniquement possible lorsque le symbiote n’est pas obligatoire)
Les manipulateurs de la reproduction chez les Arthropodes Wolbachia Rickettsia • Forte diversité • Forte incidence (> 20 % pour Wolbachia) • Peuvent atteindre des fréquences très élevées Arsenophonus Spiroplasma Cardinium Tous sont à TV! Quasiment aucun n’apporte un bénéfice
Incompatibilité cytoplasmique Féminisation Thélytoquie Male-killing n chromosomes 2n chromosomes Les manipulations de la Reproduction
Est-ce aussi simple ?TV et reproduction sexuée • Transmission uniparentale des symbiotes • Limite les infections multiples • Laisse un espace pour le conflit • Un exemple très clair que la sélection agît de manière égoïste sur chacun des partenaires !
A quelle échelle la TH est-elle importante ? Asobara tabida (1) Leptopilina heterotoma (1) 0.02 100 D. simulans (Riverside) 85 D. auraria A. albopictus 84 L. guineaensis Asobara tabida (2) 100 72 D. melanogaster C. perengrinus Pachycrepoideus dubius 100 Muscidifurax uniraptor 98 A G. centralis N. vitripennis 94 G. morsitans 88 • Pas de co-speciation • Transmission horizontale • infections multiples possibles et fréquentes Trichopria sp. (1) Leptopilina heterotoma (3) 100 E. kuehniella T. kaykai T. bourarachae Leptopilina heterotoma (2) Trichopria drosophilae 100 Asobara tabida (3) G. austeni P. papatasi L. guineaensis 100 D. sechellia 100 D. simulans (Hawai) T. deion T. kaykai T. kaykai 86 T. sibericum 98 T. nubilale T. deion 83 T. confusum T. bedeguaris 100 Trichopria sp. (2) 97 L. striatellus A. diversicornis 74 E. formosa E. cautella D. rosae S. fuscipes 99 72 T. orizicolus L. victoriae 100 L. australis L. clavipes 92 B A. albopictus C. quinquefasciatus 100 C. pipiens D. simulans (Nouméa) D. mauritiana 79 A. vulgare
Transfert Horizontal Infection perdue Invasion par manipulation de la reproduction Est-ce aussi simple ?A quelle échelle la TH est-elle importante ? • Sans rare TH, les manipulateurs de la reproduction auraient disparu !
Dedeine et al., PNAS, 2001 L’évolution du “mutualisme” ? • Le seul trait affecté (e.g. les mâles sont fertiles)
Ovogenèse Incompatibilité Cytoplasmique Incompatibilité Cytoplasmique Spécialisation des souches de Wolbachia Asobara tabida (wAtab3) Leptopilina heterotoma 100 D. simulans (Riverside) 85 D. auraria Aedes albopictus 84 Amitus fuscipennis Asobara tabida (wAtab1) 100 72 D. melanogaster Coccidoxenoides A perengrinus Pachycrepoideus dubius 100 Muscidifurax uniraptor 98 Glossina centralis Nasonia vitripennis 94 88 Glossina morsitans Trichopria sp. Leptopilina heterotoma 100 Ephestia kuehniella Trichogramma kaykai Trichogramma bourarachae Leptopilina heterotoma Trichopria drosophilae 100 Asobara tabida (wAtab2) Glossina austeni Phlebotomus papatasi Ephestia cautella 100 D. sechellia 100 D. simulans (Hawai) B Gène wsp (Wolbachia outer surface protein) Vavre et al., MBE, 1999 ; Dedeine et al., Evolution, 2004
Asobara (ITS2) 0.02 A. persimilis 100 IASPITS A. japonica AJ1 A. tabida (Saanich, USA) IATSAITS 99 A. tabida (Seattle, USA) 75 IATSEITS 83 A. tabida (Kos, Grêce) 100 IATKOITS A. rufescens IARUFITS A. citri IACIITS La dépendance a été acquise récemment ! U wAjap Dépendance wAtab1, 2 and 3 wAtab1 U Dedeine et al., Heredity, 2005
Saanick (CDN) Seattle (US) Variation intra-spécifique de l’hôte pour la dépendance 12 populations Européennes 0 (pas d’ovocyte) Infecté (W+) Non infecté (W-) 0 100 200 300 400 Charge ovarienne Dedeine et al., Heredity, 2005
Devenir des ovocytes produits Volume des ovocytes Les œufs sont pondus Ils se développent en larves Aucun adulte ne se développe La dépendance reste totale
Le cas du mutant Sxl chez Drosophila melanogaster Un seul gène impliqué ! infecté (wmel) aposymbiotique Déterminisme du phénotype ovarien La variation est contrôlée par l’hôte Charge ovarienne
Apoptose durant l’ovogenèse : que sait-on chez la Drosophile Germarium checkpoint mid-oogenesis checkpoint constitutive apoptosis Comment un hôte peut-il devenir dépendant pour une telle fonction ? Uninfected Infected Pannebakker et al., PNAS, 2007
Apoptose, immunité et pathogènes intracellulaires (e.g. Rickettsia, Anaplasma NF-KB pathway) Pathogen Dissémination dans l’organisme Destruction des cellules immunitaires - + Prolongation de la survie cellulaire APOPTOSIS + Destruction des cellules infectées Par des pathogènes intracellulaires Host
Oogenesis Apoptosis N W Compensation Oogenesis Apoptosis Apoptosis Oogenesis N W N W Wolbachia est principalement localisé dans les ovaires De la pathogénie à la dépendance ? Manipulation de l’apoptose pourrait être un phénomène général (Bazzochi et al. 2007) La dépendance est -elle une conséquence d’une stratégie parasitaire ?
Symbiotiques Aposymbiotiques Les gènes impliqués côté hôte Etude du transcriptome par banque soustractive (Suppression Subtractive Hybridization)
Résultats préliminaires SSH • Protéines ribosomales : S6, S18, L17, L23 …(e-value : 10-20 à 10-80 ) • Cyclophiline : peptidyl-propyl cis-trans isomérase(e-value : 3.10-9) • chaperonne : activité de facteurs apoptotiques (AIF) • perméabilité membranaire (mitochondrie) • Chaine lourde de laFerritine: Stockage du fer(e-value : 5.10-19) • régulé par NF-B, influence sur voie JNK (apoptose) • Fonction inconnues…
Toll pathway Imd pathway JNK pathway JAK/STAT GNBP1 NEC Persephone PGRP-SA Spätzle Mb cellulaire PGRP-LC Domeless Toll Imd MyD88 TAK1 Pelle FADD IAP2 Tube Hem Hopscotch Dredd Kenny IRD Basket Dif* Stat92E Cactus Relish* Dorsal* Jra Kay Quels gènes étudier ? • A. tabida : aucune information génétique • Séquençage de 30 000 ESTs (différentes banques) • Annotation terminée, + de 10 000 unigènes






















