Download
1 / 1
10 likes | 142 Views
SHELF LIFE OF SEEDS IS RELATED TO THE MOLECULAR MOBILITY IN THE CYTOPLASM. Julia Buitink *,# , Folkert A. Hoekstra * and Marcus A. Hemminga #

E N D
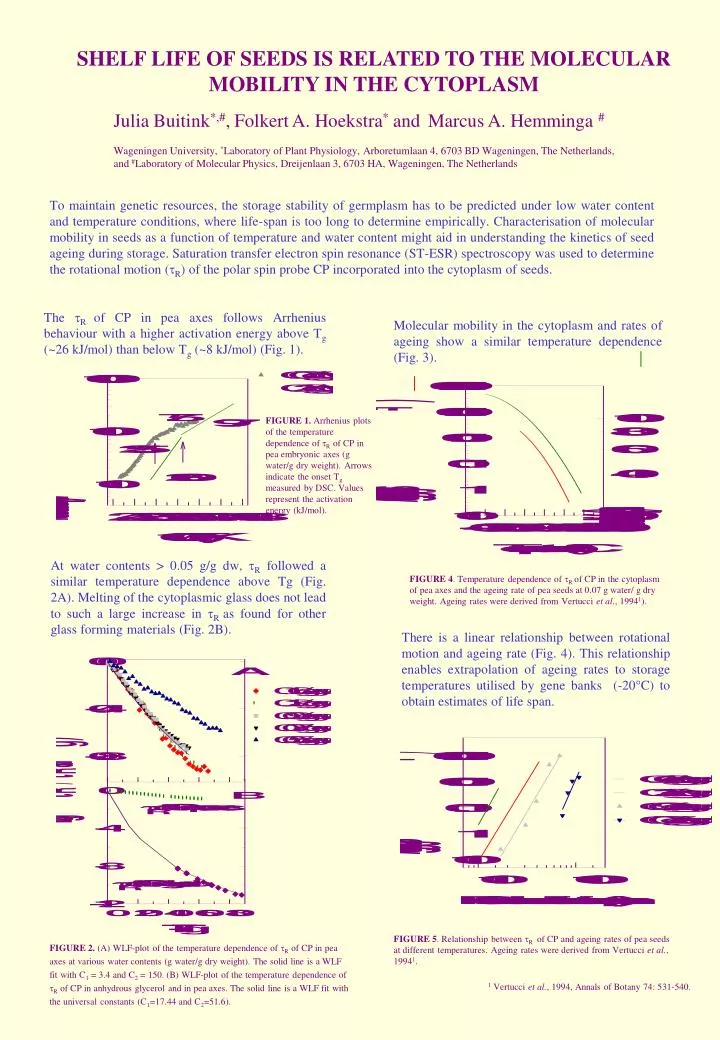
SHELF LIFE OF SEEDS IS RELATED TO THE MOLECULAR MOBILITY IN THE CYTOPLASM Julia Buitink*,#, Folkert A. Hoekstra* and Marcus A. Hemminga # Wageningen University, *Laboratory of Plant Physiology, Arboretumlaan 4, 6703 BD Wageningen, The Netherlands, and #Laboratory of Molecular Physics, Dreijenlaan 3, 6703 HA, Wageningen, The Netherlands To maintain genetic resources, the storage stability of germplasm has to be predicted under low water content and temperature conditions, where life-span is too long to determine empirically. Characterisation of molecular mobility in seeds as a function of temperature and water content might aid in understanding the kinetics of seed ageing during storage. Saturation transfer electron spin resonance (ST-ESR) spectroscopy was used to determine the rotational motion (tR) of the polar spin probe CP incorporated into the cytoplasm of seeds. FIGURE 2. (A) WLF-plot of the temperature dependence of R of CP in pea axes at various water contents (g water/g dry weight). The solid line is a WLF fit with C1 = 3.4 and C2 = 150. (B) WLF-plot of the temperature dependence of R of CP in anhydrous glycerol and in pea axes. The solid line is a WLF fit with the universal constants (C1=17.44 and C2=51.6). The R of CP in pea axes follows Arrhenius behaviour with a higher activation energy above Tg (~26 kJ/mol) than below Tg (~8 kJ/mol) (Fig. 1). Molecular mobility in the cytoplasm and rates of ageing show a similar temperature dependence (Fig. 3). FIGURE 1. Arrhenius plots of the temperature dependence of R of CP in pea embryonic axes (g water/g dry weight). Arrows indicate the onset Tg measured by DSC. Values represent the activation energy (kJ/mol). At water contents > 0.05 g/g dw, R followed a similar temperature dependence above Tg (Fig. 2A). Melting of the cytoplasmic glass does not lead to such a large increase in R as found for other glass forming materials (Fig. 2B). FIGURE 4. Temperature dependence of Rof CP in the cytoplasm of pea axes and the ageing rate of pea seeds at 0.07 g water/ g dry weight. Ageing rates were derived from Vertucci et al., 19941). There is a linear relationship between rotational motion and ageing rate (Fig. 4). This relationship enables extrapolation of ageing rates to storage temperatures utilised by gene banks (-20°C) to obtain estimates of life span. FIGURE 5. Relationship between R of CP and ageing rates of pea seeds at different temperatures. Ageing rates were derived from Vertucci et al., 19941. 1 Vertucci et al.,1994, Annals of Botany 74: 531-540.