Download
1 / 22
220 likes | 447 Views
DNA structure: Non-helical secondary structures: non-helical , palindrome, hairpin, and cruciform. Lecture 9: 1 10/11/2006.

E N D
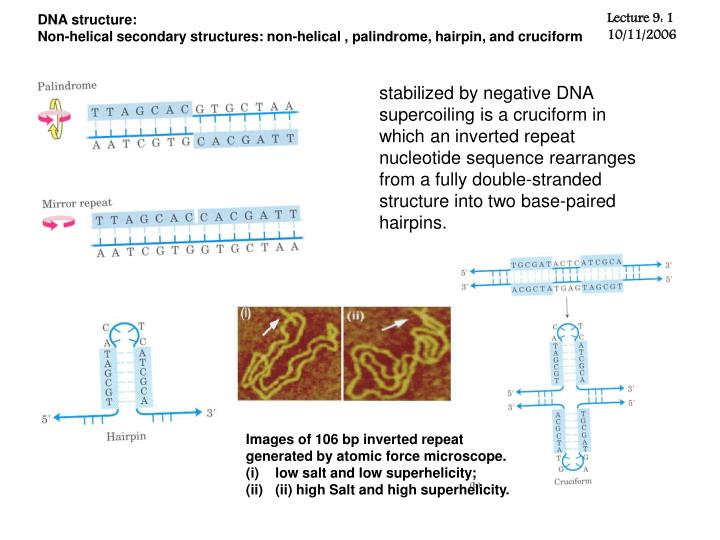
DNA structure: Non-helical secondary structures: non-helical , palindrome, hairpin, and cruciform Lecture 9: 1 10/11/2006 stabilized by negative DNA supercoiling is a cruciform in which an inverted repeat nucleotide sequence rearranges from a fully double-stranded structure into two base-paired hairpins. • Images of 106 bp inverted repeat • generated by atomic force microscope. • low salt and low superhelicity; • (ii) high Salt and high superhelicity.
DNA structure: Tertiary Structure: Plasmid Lecture 8: 2 10/11/2006 Primary features of plasmids: 1) extrachromosomal DNA; 2) capable of autonomous replication. • Some other common features: • It is typically circular and double-stranded. It usually occurs in bacteria, sometimes in eukaryotic organisms (e.g., the 2-micrometre-ring in Saccharomyces cerevisiae). • Size of plasmids varies from 1 to over 400 kilobase pairs (kbp). • There may be one copy, for large plasmids, to hundreds of copies of the same plasmid in a single cell, or even thousands of copies. stringent - low copy number (F factor); relaxed - high copy number (pBR322 - 16 copies; pUC - 30 to 50 copies) • Some plasmids carry antibiotic resistance gene (s) or reporter genes. • 5. Multiple cloning sites are present in commercial plasmid vectors. • 6. The term plasmid was first introduced by the American molecular biologist Joshua Lederberg in 1952.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 3 10/11/2006 TEM images of various Forms of a plasmid Negative and positive superhelical tension occur when the helix is unwound or over wound, respectively In the cells, plasmids are negatively supercoiled, thus favoring the melting and interactions with proteins.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 4 10/11/2006
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 5 10/11/2006 Basics of Optics the relation between focal length f, object distance a, and image distance b which is called the lens equation: 1 / a + 1 / b = 1 / f Lens aberrations (a) spherical aberration, (b) chromatic aberration, (c) astigmatism, and (d) coma.
The distance from the center of the convex lens to the focal plane is know as the focal distance. (For an idealized symmetrical thin convex lens, this distance is the same in front of or behind the lens.) The image of our giraffe now appears at the focal plane (as illustrated in Figure 2). The image is smaller than the object (the giraffe); it is inverted and is a real image capable of being captured on film. This is the case for the camera used for ordinary scenic photography. The object is now moved closer to the front of the lens but is still more than two focal lengths in front of the lens (this scenario is addressed in Figure 3). Now, the image is found further behind the lens. It is larger than the one described above, but is still smaller than the object. The image is inverted, and is a real image. This is the case for ordinary portrait photography. The object is brought to twice the focal distance in front of the lens. The image is now two focal lengths behind the lens as illustrated in Figure 4. It is the same size as the object; it is real and inverted. The object is now situated between one and two focal lengths in front of the lens (shown in Figure 5). Now the image is still further away from the back of the lens. This time, the image is magnified and is larger than the object; it is still inverted and it is real. This case describes the functioning of all finite tube length objectives used in microscopy. Such finite tube length objectives project a real, inverted, and magnified image into the body tube of the microscope. This image comes into focus at the plane of the fixed diaphragm in the eyepiece. The distance from the back focal plane of the objective (not necessarily its back lens) to the plane of the fixed diaphragm of the eyepiece is known as the optical tube length of the objective.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 7 10/11/2006 Basics of Optics In the last case, the object is situated at the front focal plane of the convex lens. In this case, the rays of light emerge from the lens in parallel. The image is located on the same side of the lens as the object, and it appears upright (see Figure 1). The image is a virtual image and appears as if it were 10 inches from the eye, similar to the functioning of a simple magnifying glass; the magnification factor depends on the curvature of the lens. The last case listed above describes the functioning of the observation eyepiece of the microscope. The "object" examined by the eyepiece is the magnified, inverted, real image projected by the objective. When the human eye is placed above the eyepiece, the lens and cornea of the eye "look" at this secondarily magnified virtual image and see this virtual image as if it were 10 inches from the eye, near the base of the microscope. Since the image appears to be on the same side of the lens as the object, it cannot be projected onto a screen. Such images are termed virtual images and they appear upright, not inverted. Figure 1 presents an illustration of how a simple magnifying lens operates. The object (in this case the subject is a rose) is being viewed with a simple bi-convex lens. Light reflected from the rose enters the lens in straight lines as illustrated in Figure 1. This light is refracted and focused by the lens to produce a virtual image on the retina. The image of the rose is magnified because we perceive the actual size of the object (the rose) to be at infinity because our eyes trace the light rays back in straight lines to the virtual image (Figure 1).
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 8 10/11/2006 Basics of Optics Multi-lens imaging systems achieve very high magnification.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 9 10/11/2006 Bragg's Law can easily be derived by considering the conditions necessary to make the phases of the beams coincide when the incident angle equals and reflecting angle. The rays of the incident beam are always in phase and parallel up to the point at which the top beam strikes the top layer at atom z (Fig. 1). The second beam continues to the next layer where it is scattered by atom B. The second beam must travel the extra distance AB + BC if the two beams are to continue traveling adjacent and parallel. This extra distance must be an integral (n) multiple of the wavelength (l) for the phases of the two beams to be the same: nλ = AB +BC (2). Bragg's Law refers to the simple equation: nλ = 2d sinθ Fig. 1 Deriving Bragg's Law using the reflection geometry and applying trigonometry. The lower beam must travel the extra distance (AB + BC) to continue traveling parallel and adjacent to the top beam. Recognizing d as the hypotenuse of the right triangle Abz, we can use trigonometry to relate d and θ to the distance (AB + BC). The distance AB is opposite q so, AB = d sinθ (3). Because AB = BC eq. (2) becomes, nλ = 2AB (4) Substituting eq. (3) in eq. (4) we have, nl = 2 d sinθ, (1) and Bragg's Law has been derived. The location of the surface does not change the derivation of Bragg's Law.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 10 10/11/2006 Microscopy The resolution of an optical microscope is defined as the shortest distance between two points on a specimen that can still be distinguished by the observer or camera system as separate entities. There are several equations that have been derived to express the relationship between numerical aperture, wavelength, and resolution: Resolution (r) = λ/(2NA) (1) Resolution (r) = 0.61λ/NA (2) Resolution (r) = 1.22λ/(NA(obj) + NA(cond)) (3) Where r is resolution (the smallest resolvable distance between two objects), NA is a general term for the microscope numerical aperture, is the imaging wavelength, NA(obj) equals the objective numerical aperture, and NA(cond) is the condenser numerical aperture. Notice that equation (1) and (2) differ by the multiplication factor, which is 0.5 for equation (1) and 0.61 for equation (2).
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 11 10/11/2006 Microscopy Numerical Aperture Numerical Aperture (also termed Object-Side Aperture) is a value (often symbolized by the abbreviation NA) originally defined by Abbe for microscope objectives and condensers. It is given by the simple expression: Numerical Aperture (NA) = n • sin(α ) or n • sin(θ) Note: Many authors use the variable m to designate the one-half angular aperture while others employ the more common term α , and in some instances, θ. In the numerical aperture equation, n represents the refractive index of the medium between the objective front lens and the specimen, and m or a is the one-half angular aperture of the objective.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 12 10/11/2006 Microscopy Objective types Plan Achromat Plan Fluorite Plan Apochromat Magnifi N.A. R N.A. R N.A. R 4x 0.10 2.75 0.13 2.12 0.20 1.375 10x 0.25 1.10 0.30 0.92 0.45 0.61 20x 0.40 0.69 0.50 0.55 0.75 0.37 40x 0.65 0.42 0.75 0.37 0.95 0.29 60x 0.75 0.37 0.85 0.32 0.95 0.29 100x 1.25 0.22 1.30 0.21 1.40 0.20 N.A. = Numerical Aperture R=reolution (microm) Wavelength (nm) Resolution(μM) 360 .19 400 .21 450 .24 500 .26 550 .29 600 .32 650 .34 700 .37
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 13 10/11/2006 Microscopy
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 14 10/11/2006 Basics of magnetic lens A strong rotationally symmetric, magnetic field between a pair of cylindrical pole pieces drives paraxially moving particles along a spiral path that converges towards the axis. This results from magnetic field components perpendicular as well as parallel to the optical axis (Fig.a). Thus, a magnetic field of this kind serves as a lens for charged particle beams. Rotationally symmetric electrostatic fields are created between cylindrical electrodes. Trajec tories of charged particles are bent as a result of the electric field and converge towards a point on the optical axis. (Fig. b).
objective projective objective DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 15 10/11/2006 Comparison of two types of electron microscopes
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 16 10/11/2006 Negative staining of biological molecules for electron microscopy 1) Macromolecules or supramolecular assemblies are adsorbed from their suspension onto a thin carbon support film; 2. Washed with an aqueous solution of a heavy metal salt (e.g., 1% uranyl acetate or 2% Na-phosphotungstate), before the sample is allowed to dry. A very thin film of metal salt now covers the support film everywhere except where it has been excluded by the presence of an adsorbed macromolecule. 3. Observe under TEV. Because the macromolecule allows the electrons to pass much more readily than does the surrounding heavy metal film, a reversed or negative image of the molecule is created, hence the name “negative staining”
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 17 10/11/2006 Agarose Gel Analysis of Biological Molecules (e. g. DNA)
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 18 10/11/2006 Chemical and physical properties of agarose Agar is an unbranched polysaccharide obtained from the cell walls of some species of red algae or seaweed. The word agar comes from the Malay word agar-agar (meaning jelly). It is also known as kanten or agal-agal (Ceylon agar). Chemically, agar is a polymer made up of subunits of the sugar galactose. Agar polysaccharides serve as the primary structural support for the algae's cell walls. Dissolved in hot water and cooled, agar becomes gelatinous. Its chief use is as a culture medium for microbiological work. Other uses are as a laxative, a vegetariangelatin substitute, a thickener for soups, in jellies, ice cream and Japanese desserts such as anmitsu, as a clarifying agent in brewing, and for paper sizing fabrics. Typical Properties- Gelling temp : 26º-30ºC - Melting temp : ≤65ºC - Moisture content : ≤10% - Sulfate : ≤0.10% - EEO, (-mr) : ≤0.10 - Gel strength : ≥200 g/cm2 - RNase/Dnase Activity : None Detected
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 19 10/11/2006 G. parvispora or G. tikvahiae Properties of Agarose Source of agarose Agarose is derived from a series of naturally occurring derivatives from seaweed. Most agar comes from various species of Gelidium and Gracilaria. All species contain ester sulfates and some, except Gracilaria, contain varying amounts of pyruvates. Gracilaria agarose contains methyl ethers, the position of which is variable according to the species. Structure of agarose Agarose consists of 1,3-linked ß-D-galactopyranose and 1,4-linked 3,6-anhydro-α-L-galactopyranose. This basic agarobiose repeat unit forms long chains with an average molecular mass of 120,000 daltons, representing about 400 agarobiose units. There are also charged groups present on the polysaccharide, most notably pyruvates and sulfates.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 20 10/11/2006 Properties of Agarose Electroendosmosis (EEO) Electroendosmosis (EEO) is a functional measure of the number of sulfate and pyruvate residues present on the agarose polysaccharide. This phenomenon occurs during electrophoresis when the anticonvective medium (the agarose in this case) has a fixed negative charge. In an electric field, the hydrated positive ions associated with the fixed anionic groups in the agarose gel migrate toward the cathode. Water is thus pulled along with the positive ions, and migration of negative molecules such as DNA is retarded. How EEO is measured Electroendosmosis is quantitated by subjecting a mixture of dextran and albumin to electrophoresis, then visualizing them and measuring their respective distances from the origin. The amount of EEO expressed in terms of relative mobility (-mr) is calculated by dividing the migration distance of the neutral dextran (OD) by the sum of the migration distances of the dextran and the albumin (OD + OA): -mr = OD/(OD + OA).
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 21 10/11/2006 Properties of Agarose Gelation The mechanism for gelation of agarose was first suggested by Rees and later demonstrated by Arnott. It involves a shift from a random coil in solution to a double helix in the initial stages of gelation, and then to bundles of double helices at the final stage. The average pore size varies with concentration and type of agarose, but is typically 100 to 300 nm.
DNA structure: Tertiary Structure: topology of plasmids Lecture 7: 22 10/11/2006 Properties of Agarose Methylation of agarose The agarose polysaccharide also contains uncharged methyl groups. The extent of natural methylation is directly proportional to the gelling temperature. Unexpectedly, synthetically methylated agaroses have lower, rather than higher, gelling temperatures, and the degree of synthetic methylation is inversely proportional to the gelling temperature.


















