Download
1 / 18
240 likes | 774 Views
Chapter 9 DNA-Protein Interactions in Bacteria. Student learning outcomes: Describe examples of structure /function relationships in phage repressors Appreciate that altered specificity repressors and operator mutants clarify mechanisms of amino acid: base pair recognition

E N D
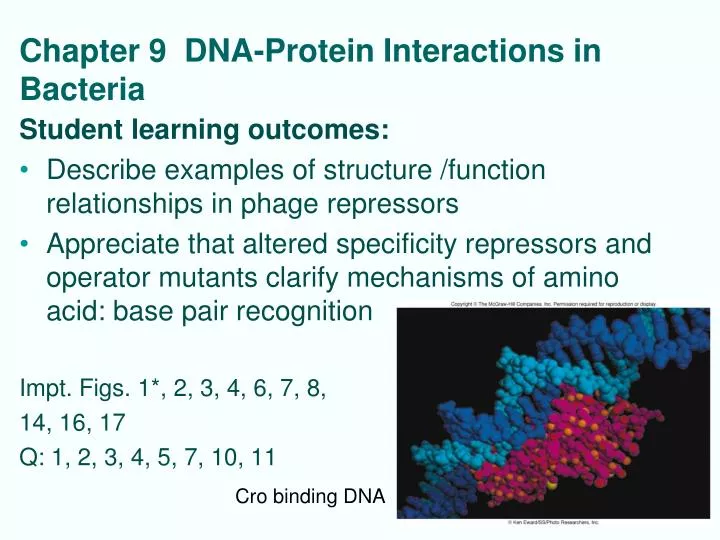
Chapter 9 DNA-Protein Interactions in Bacteria Student learning outcomes: • Describe examples of structure /function relationships in phage repressors • Appreciate that altered specificity repressors and operator mutants clarify mechanisms of amino acid: base pair recognition Impt. Figs. 1*, 2, 3, 4, 6, 7, 8, 14, 16, 17 Q: 1, 2, 3, 4, 5, 7, 10, 11 Cro binding DNA
The l Family of Repressors: l, 434, P22 • Repressors have recognition helices that lie in major groove of appropriate operator • Helix-turn-helix motif (HTH) • Specificity of bp binding depends on amino acids in recognition helices • Phages are not immune to super-infection by each other Fig. 2
Binding Specificity of lambda-like Repressor: Operator DNA • Recognition helices fit sideways in major groove of operator DNA • Certain amino acids on DNA side of recognition helix 2 make specific contact with bases in operator • Contacts determine specificity of protein-DNA binding • ** Changing amino acids can change specificity of repressor to different DNA sequence Fig. 1
Probing Binding Specificity by Site-Directed Mutagenesis - Mark Ptashne Key amino acids in recognition helices of P22, 434 repressors proposed Amino acids differ between repressorss Change 5 aa of 434 to P22; see altered specificity repressor binds P22 DNA Fig. 4 DNase footprint Fig. 3
l Repressor • l repressor has extra motif, N-terminal arm that aids binding by embracing DNA • Cro and l repressors share affinity for same operators, but micro-specificities for OR1(l) or OR3 (cro) • Specificities determined by interactions between different amino acids in recognition helices and different base pairs in operators Fig. 4 l repressor dimer on OR2
High-Resolution Analysis of lRepressor-Operator co-crystal • Recognition helices (3 red) of each monomer nestle into DNA major grooves (2 half-sites) • Helices hold two monomers together in repressor dimer • DNA is similar to B-form DNA • DNA bends at ends of fragment as curves around repressor dimer Fig. 6; operator sequence; Fig. 7 model
Amino acids of l repressor hydrogen bond with Bases in major groove Fig. 8
Amino Acid: DNA Backbone Interactions • Hydrogen bond at Gln33 maximizes electrostatic attraction between positively charged amino end of a-helix and negatively charged DNA • Attraction works to stabilize bond Fig. 9
High-Resolution Analysis of 434 Repressor-Operator Interactions • Genetic and biochemical data predicted R-O contacts • X-ray crystallography of 434 repressor-fragment/ operator-fragment shows H bonding at Gln residues in recognition helix to 3 bp in DNA • Potential van der Waals contact between Gln29 and 5Me of T3 Fig. 10
Phage 434: Effects on DNA Conformation • R-O complex DNA deviates from normal shape • DNA bends to accommodate base /aa contacts • Central part of helix is wound extra tightly • Outer parts are wound more loosely than normal • DNA sequence of operator facilitates bending Fig. 11 Normal DNA; DNA bent by 434 repressor binding
9.2 trp Repressor and role of Tryptophan • trp repressor uses helix-turn-helix (HTH) DNA binding motif to contact operator • Aporepressor is not active in binding DNA • Tryptophan forces recognition helices of trp repressor dimer into proper position to bind trp operator Fig. 12
9.3 General Considerations on Protein-DNA Interactions; multimeric proteins • Specificity of binding between protein and specific stretch of DNA relates to: • Specific interactions between bases and amino acids • Ability of DNA to assume shape that directly relates to DNA’s base sequence • Target sites for DNA-binding proteins usually symmetric or repeated • Most DNA-binding proteins are dimers: greatly enhances binding between DNA and protein as protein subunits bind cooperatively
Hydrogen Bonding Capabilities of Different Base Pairs • Protein ‘reads the DNA’ • Different base pairs present four different hydrogen-bonding profiles to amino acids approaching either major or minor groove Fig. 14
9.4 DNA-Binding Proteins: Action at a Distance • DNA-binding proteins can influence interactions at remote sites in DNA – often looping intervening DNA • Common in eukaryotes • Occurs in several prokaryote systems: lac operon multiple operators ara operon looping gal operon looping l repressor
E. coli gal Operon • gal operon has 2 operators, 97 bp apart • One adjacent to gal promoter - External operator, OE • Other located within first structural gene, galE Oi • 2 separated operators - both bind repressors that interact, loop out intervening DNA • Recall Chapt. 7 lacI, araC repressors Fig. 15
DNA Looping affects DNase Susceptibility Fig. 16 Operators separated by • Integral number of double-helical turns loop out DNA to allow cooperative binding • Nonintegral number of turns requires proteins to bind opposite faces of DNA, no cooperative binding Fig. 17 l repressor binds cooperatively to operators
Enhancers Enhancers are nonpromoter DNA elements that bind protein factors and stimulate transcription • Can act at a distance • Originally found in eukaryotes (lots chapt. 12) • Recently found in prokaryotes. E. coli glnA gene: • NtrC protein binds enhancer, • Binds RNAP 70 bp away • NtrC hydrolyzes ATP, lets RPo form. • Insert 350 bp, see loop Fig. 20 NtrC: RNAP
Review questions • 1. Draw rough diagram of helix-turn-helix domain interacting with DNA double helix • 2. Describe experiment that shows which amino acids bind which base pairs in l-like phage repressors. • 10. Explain fact that protein oligomers (dimers, tetramers) bind better to DNA than monomeric proteins