Download

1 / 75
760 likes | 973 Views
The trp operon. Suggested readings on regulation/dna bp Voet pp 1237-1253 Problems 2, 4. Figure 31-39 A genetic map of the E. coli trp operon indicating the enzymes it specifies and the reactions they catalyze. MVA Fig.21.13. MVA Fig.21.14.

E N D
Suggested readings on regulation/dna bp Voet pp 1237-1253 Problems 2, 4
Figure 31-39 A genetic map of the E. coli trp operon indicating the enzymes it specifies and the reactions they catalyze.
Figure 31-31 X-Ray structure of an E. coli trp repressor– operator complex. Page 1244
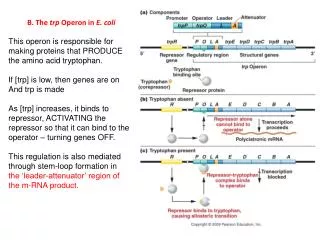
The Tryptophan Operon: A Repressor • When should the bacteria be transcribing genes for the synthesis of the amino acid tryptophan? • When levels of tryptophan in the cell are low, the bacteria must make its own. • However, if tryptophan is abundant in the cell or is being provided in the medium, it is a waste of energy for the bacteria to be synthesizing it.
Figure 31-40 The base sequence of the trp operator. The nearly palindromic sequence is boxed and its –10 region is overscored.
Originally, regulation of the trp operon was thought to occur solely through the repressor-operator system until deletion mutants located downstream of trpO were identified. These mutants displayed increased expression of the operon by six-fold which indicated the presence of an additional transcriptional control element. Why is repression not the only mode of regulation?
Evidence: • 1. Trp-tRNA synthetase mutants had regulatory anomalies. • 2. Addition of trp to trp-starved cells not only shut down • initiation of transcription but also inhibited transcription • already in progress on the initial segment of the operon • 3. Mutants lacking a functional repressor could still respond to trp starvation by increasing transcription of trp mRNA. • 4 . Deletion mutants in which both of the deletion termini were within the transcribed region of the operon had an • unexpected six-fold increase in expression of the remaining • genes in the operon. Obviously, repressor binding was • unaffected.
5. Within the population of mRNAs produced in vivo from the 5' end of the trp operon, RNAs corresponding to the first 140 bp (the leader sequence) of the operon were several times more abundant than those from more distal regions, therefore a transcription termination site was located before the structural genes. 6. Starving bacteria of trp reduced termination at this site (the trp attenuator). 7. Mutations altering trp-tRNA synthetase, tRNAtrp or a tRNA trp modifying enzyme were found to decrease transcription termination at the trp attenuator. What does this suggest about the mode of attenuation?
8. Ribosome binding experiments with the 140 base transcript • demonstrated that ribosomes protect a 20 base segment from • nuclease attack. A potential AUG start codon is located in the • center of this region. • 9. A 14 residue peptide (the leader peptide) could be • synthesized from this start codon and contained tandem trp • residues near its C-terminus. • 10. The trp leader ribosome binding site was shown to be an • efficient site for the initiation of translation by fusing the leader • to a structural gene and demonstrating synthesis of the fused • polypeptide. Examples?
11. Two classes of termination defective leader mutants have been isolated. One type terminates at less than normal frequency and has bp changes in the 3:4 bp region. In vivo, these mutants have a 2-4 fold increase in operon expression. • 12. The second class of mutants have increased termination of • the attenuator. These prevent the relief from termination that • is associated with trp starvation. One of these mutants has an • altered start codon for the leader peptide. Another has a • G to A conversion at position 75, which would prevent • 2:3 pairing and cause formation of a 3:4 termination structure.
Genetic analysis indicated that the new control element was located in trpL, a 162 nt region 30-60 nt upstream from trpE. When trp is scarce, the entire 6720 nt polycistronic trp, including trpL, is synthesized. As the trp concentration increases, the rate of trp transcription decreases as a result of the trp r epressor-corepressor's greater abundance. With increasing [trp], the mRNA synthesized consists more and more of a 140 nt segment corresponding to trpL sequences only. The availability of trp results in the premature termination of transcription of the operon.
Figure 31-41 The alternative secondary structures of trpL mRNA. Page 1252
Figure 31-42a Attenuation in the trp operon. (a) When tryptophanyl–tRNATrp is abundant, the ribosome translates trpL mRNA.
Figure 31-42b Attenuation in the trp operon. (b) When tryptophanyl–tRNATrp is scarce, the ribosome stalls on the tandem Trp codons of segment 1.
Trp operon And again all the cool animation files
DNA Binding Proteins How does a repressor find its operator in a sea of other sequences? It is not enough just for the regulatory protein to recognize the correct DNA site. The protein must also find it rapidly and bind to it sufficiently tightly to discriminate it from the millions of competing and overlapping nonspecific sites that are explored in the course of specific target localization.
One point to keep in mind while considering protein-DNA interactions is that such an interaction represents a dynamic equilibrium: Whether an operator has its (or a) particular repressor protein bound to it depends on: 1. the concentration of the regulatory protein in the cell, 2. the affinity between the repressor and the operator sequence and 3. the affinity between the repressor and other non-specific DNA binding sites.
Association constants: lac repressor + DNA to R-DNA complex Repressor: lac operator 1-2 X 1013 M-1 other DNA 2-3 X 106 M-1 (specificity = KA(s)/KA(non-specific) = 107) Repressor bound to inducer lac operator 2 X 1010 M-1--or some references suggest this is even lower other DNA 2 X 106 M-1 When repressor is bound to allosteric regulator (allolactose in this case) non-specific binding competes more effectively with specific binding.
How a repressor recognizes and binds to an operator The interaction between repressor and operator is often taken as a paradigm for sequence-specific DNA-protein interactions. Each regulatory protein in E. coli must select its operator site (or sites) from among the five million or so base pairs of DNA in the cell. Examples? For this organism, an operator (or any other cis acting site) must be at least 11-12 bases long in order to form a site that reoccurs at random less than once per genome. Accordingly, regulatory proteins in E. coli bind tightly to specific DNA sequences that are about 15-20 base pairs long.
Operator Sequence and Structure: A large number of operator sites have been identified and their DNA sequence has been determined. One feature that is common to all operators is an imperfect two-fold axis of symmetry. • A perfectly symmetrical sequence is shown below. • >---- G C C A T G C G C A T G G C ----> • <---- C G G T A C G C G T A C C G ----<
Cap binding site: Link to view structure
Lac repressor lac operator: binding site for the lac repressor protein (lac I gene product)
Structure of Regulatory Proteins: Many DNA-binding regulatory proteins share features in common that reflect a common mode of DNA binding. Some of these features are: (1) The active binding unit is a dimer of two identical globular polypeptide chains oriented oppositely in space to give a molecule with a two-fold axis of symmetry phage lambda cI repressor protein alpha helical region in contact with the major groove is in red.
(2) The critical contacts between the protein and the DNA are made by adjacent a helices located at the binding face of each monomer. The helices are connected by a turn in the protein secondary structure. This helix-turn-helix motif is common to many regulatory proteins. HTH
Cro Why do the recognition helices contact the major groove? What determines the specificity of interaction?
Binding motifs http://www.umass.edu/molvis/freichsman/Site642/page_dnab/menu.html again http://www.web-books.com/MoBio/Free/Ch4F2.htm#4F1
A=H bond acceptor D=H bond donor
One of the most common DNA-protein interactions. Because of its specific geometry of H-bond acceptors, guanine can be unambiguously recognized by the side chain of arginine
Specificity of protein-DNA interaction of due to: • ability of amino acid side chains in the recognition helix to form hydrogen bonds with specific bases in its cis-acting site • multiple complementary interactions between the protein and the DNA that are dependent on the deformation of the helix and which increase the number of contact points
Main features of interactions between DNA and the helix-turn-helix motif of DNA binding proteins