Download
1 / 73
730 likes | 1.22k Views
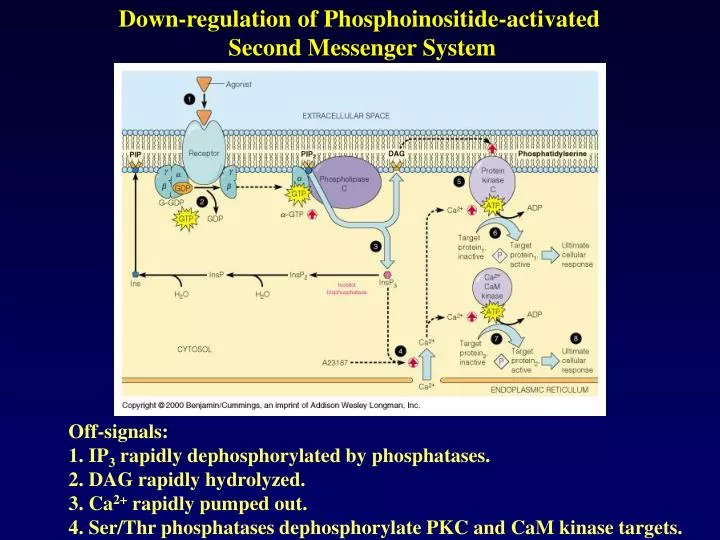
Down-regulation of Phosphoinositide-activated Second Messenger System. Off-signals: 1. IP 3 rapidly dephosphorylated by phosphatases. 2. DAG rapidly hydrolyzed. 3. Ca 2+ rapidly pumped out. 4. Ser/Thr phosphatases dephosphorylate PKC and CaM kinase targets.

E N D
Down-regulation of Phosphoinositide-activated Second Messenger System Off-signals: 1. IP3 rapidly dephosphorylated by phosphatases. 2. DAG rapidly hydrolyzed. 3. Ca2+ rapidly pumped out. 4. Ser/Thr phosphatases dephosphorylate PKC and CaM kinase targets.
Fertilization of an Egg by a Sperm Triggering an Increase in Cytosolic Ca2+ Prior to fertilization, the starfish egg was injected with a Ca2+-sensitive fluorescent dye. A wave of cytosolic Ca2+ (red), released from the ER, is seen to sweep across the egg from the site of sperm entry.
Wave of Ca2+ release and PKC activation spreading from the site of artificial activation of a Xenopus egg. The egg was injected with calcium red (a fluorescent dye sensitive to the [Ca2+]) and a fusion protein, consisting of green fluorescent protein and PKC, which produces green fluorescence when PKC is activated.
Protease-Activated Receptors (PARs): GPCR’s activated by a novel proteolytic mechanism: Protease forms a Michaelis complex with its receptor/substrate (E•S) to proteolytically cleave a small peptide from the NH2-terminus generating a “NEW” NH2-terminus This “new” NH2-terminus serves as a tethered peptide ligand, binding intramolec- ularly to the body of the “receptor” to effect transmembrane signaling. Proteases are typically serine proteases. Platelet Thrombin receptor structure: PAR1 Mechanisms of Platelet Activation and Control, Plenum Press, 1993
Features of PAR Amino-terminal Exodomains PARs’ hirudin-like sequences Hirudin C-tail DFEEIPEEYLQ PAR1 38 L-DPR // SFLLR-NPNDKYEPFWEDEEK 61 PAR3 35 L-PIK // TFRGA-PPN-SFEEFPFSALE 57 PAR4 43 LPAPR // GYPGQVCANDSDTLELPDSSR 68 PAR2 33 S-KGR // SLIGK-VDGTSHVTGKGVTV- 55 Cleavage site Tethered ligand domains
Cleavage of the receptor/substrate by the protease results in “Irreversible” activation Irreversible Activation by Cleavage “Reversible” activation can be accomplished with a small peptide mimicking the new NH2-terminus Reversible Activation by Peptide Mimetic
One Receptor Can Couple to More than One G protein Four different receptors (R1-4) are shown. R3 couples to both Gi and Gq
Modulation of Glycogen Synthase Activity by Phosphorylation This enzyme contains nine separate sites in five designated regions susceptible to phosphorylation by several different cellular protein kinases. Consequently, the activity of this enzyme can be modulated in response to a variety of second messengers produced in response to different extracellular signals. Therefore regulation is a matter of finely tuned modulation of the enzyme activity over a wide range and not merely on/off switching.
Structural/functional Consequences of Lipid Modifications TIBS 16:338, 1991 • Part of the GPCR C-terminus is tethered to the membrane by palmitoylation through a thioester linkage to a conserved Cys. • The G protein subunit has myristate attached via an amide linkage to an N-terminal Gly to effect membrane association. • The G protein subunit is incorporated into the membrane via a geranylgeranyl moiety attached via a thioether linkage to • a C-terminal conserved Cys. • Some G protein-coupled effectors are transmembrane proteins (e.g. ion channels and adenylyl cyclase) as shown here, • others (e.g. phospholipase C) may be peripheral membrane proteins.
Sites of Lipid Modification of G Protein Subunits TIBS 16:338, 1991
Production of the 2nd Messenger – cGMP The membrane-bound form of guanylate cyclase is acti- vated directly by hormone binding to its receptor, e.g. atrial natriuretic peptides. The cytosolic form is a different protein and is activated by nitric oxide (NO). Protein kinase G (PKG) is activated by cGMP.
Functional Domains of the ANF Receptor Figure 21.44 – Devlin, Textbook of Biochemistry
I. Cell Signaling by Nitric Oxide (NO) Most cells can produce NO; however, it appears to function primarily in three broad categories: 1. via the endothelial cell to cause vascular (smooth muscle cell) relaxation; 2. during neurotransmission to facilitate CNS function; 3. in cell-mediated immune responses to facilitate immunologic function. NO is produced via the enzyme complex, NO synthase (NOS), which deaminates Arg to form citrulline and NO
II. Cell Signalling by NO Basal production of NO required to control vascular tone and normal blood pressure. NO is chemically unstable and decomposes in seconds!!! NO is chemically unstable and decomposes in seconds!!! NO-mediated intercellular communication: Agonists such as acetylcholine, thrombin, histamine and bradykinin activate their various receptors on vascular endothelial cells, leading to activation of NO synthase. NO diffuses from the EC to the smooth muscle cell (SMC) Within the SMC, NO stimulates production of cGMP via activation of a soluble guanylate cyclase. cGMP activates cGMP-dependent kinases (PKG) with the end result being SMC relaxation. Nitroglycerin and nitroprusside, well known for treating angina pectoris function by producing NO during their metabolism.
Second Messengers Involved in Intracellular Signaling © 2000 by W. H. Freeman and Company. All rights reserved.
Eukaryotic Signal Transduction Systems Involving Membrane Receptors (1-5) and Second Messengers (1-4)
Second Messengers Involved in Intracellular Signaling © 2000 by W. H. Freeman and Company. All rights reserved.
Signaling via Enzyme-linked Cell Surface Receptors Receptor Protein Tyrosine Kinases (RPTKs) Receptors are characterized by : 1) a single membrane-spanning domain; 2) a large glycosylated extracellular domain defining the ligand binding site; and 3) an intrinsic enzymatic activity contained within the cytoplasmic domain (tyrosine kinase activity)
Families of Ligands for Enzyme-linked Receptors Molecular Basis of Medical Cell Biology, 1998 Ligands for RPTKs
BASIC PRINCIPLES OF RPTK SIGNALING Ligand-induced RPTK Dimerization and Subsequent Phosphorylation © 2000 by W. H. Freeman and Company. All rights reserved. Some ligands are monomeric, e.g. EGF. Ligand binding induces a conformational change in the receptors receptor dimerization. Other ligands are dimeric and bring two receptors together directly. Irrespective, subsequent to dimerization, the kinase activity of each subunit cross- phosphorylates Tyr residues near the active site in the other subunit. Autophos- phorylation of Tyr residues within other parts of the cytoplasmic domain ensues.
BASIC PRINCIPLES OF RPTK SIGNALING Function of Phosphorylated Tyrosines © 2000 by W. H. Freeman and Company. All rights reserved. Phosphotyrosines of dimerized receptor serve as high affinity binding sites for “docking” and/or “activation” of intracellular signalling proteins.
BASIC PRINCIPLES OF RPTK SIGNALING Selected Intracellular Substrates and Targets of RPTKs Proteins that bind to Tyr-PO4 residues must contain SH2 or SH3 domains [Src(soluble cytoplasmic tyrosine kinase) Homology domains] • SH2 - 1– aa stretch; SH3- 60 aa stretch • Crystallographic studies demonstrated that these domains • have a pocket into which a Tyr-PO4 residue would fit • Adapter proteins link other cytoplasmic proteins to the • activated receptor Molecular Basis of Medical Cell Biology, 1998
Binding of Various Signal-transducing Molecules to Different Phosphorylated Tyrosines in the PDGF receptor ** **denote Tyr residues that are phosphorylated ***the outlined area in each bound signal trans- duction molecule repre- sents the SH2 domain ***
Binding pocket for a specific amino acid side chain Binding pocket for phosphotyrosine The SH2 Domain: A Compact “Plug-in” Molecule The 3-D Structure of an SH2 Domain
Phospholipase C is Coupled to Gq Phospholipase C Binds to Phosphorylated RPTKs
Mechanism of Activation of PLC-1 by EGF or PDGF • Binding of EGF (or PDGF) to its receptor elicits autophosphorylation subsequent to • receptor dimerization. (b) The PLC-1 forms a complex with the EGF (PDGF) phosphorylated • receptor and the tyrosine residues 771, 783 and 1254 are phosphorylated, which (c) activates • PLC-1.. (TIBS 16:297, 1991)
PLC-, -, - Share Regions of Significant Homology (X and Y) (G protein activated) (Tyrosine-kinase activated) (TIBS 16:297, 1991) (TIBS 16:297, 1991) TIBS 16:297, 1991
How Do Growth Factor Receptors Lead to Cell Proliferation? The Activation of Ras by an Activated RPTK The Grb-2 adaptor protein binds to a specific phosphotyrosine on the receptor and to the Ras guanine nucleotide exchange factor (GEF), which stimulates Ras to exchange its bound GDP for GTP
What are Ras proteins and how are they activated? GEF Molecular Biology of the Cell, 2002 Ras proteins are monomeric GTPases. GEFs activate Ras by stimulating it to give up its GDP for GTP. The cytosolic [GTP] is 10 times > [GDP], so Ras readily binds GTP once GDP has been ejected. Ras-GAPs inactivate Ras by stimulating its GTPase activity and thus its hydrolysis of GTP to GDP. The Ras-GAPs maintain most of the Ras protein (95%) in an unstimulated state.
** © 2000 by W. H. Freeman and Company. All rights reserved. © 2000 by W. H. Freeman and Company. All rights reserved. © 2000 by W. H. Freeman and Company. All rights reserved. Conformational Changes Accompanying Ras Activation **Sos is a GEF specific for Ras
The MAP-kinase Ser/Thr phosphorylation pathway activated by Ras MAP; mitogen activated protein Raf Mek Erk Ras activates MAP-KKK (Raf), which in turn activates MAP-KK (Mek) by phosphoryla- tion of select Ser/Thr residues. Activated Mek activates MAP-K (Erk) in an analogous manner. Erk in turn phosphorylates a variety of downstream proteins resulting in changes in gene expression and protein activity causing complex changes in cell behavior.
Growth Factor Activation of its RPTK Leading to Cell Proliferation Leading to proliferation
Insulin-dependent stimulation of glucose uptake occurs through a Ras-independent pathway! IRS binds/stimulates PI-3 kinase, which effects an, as yet, ill-defined cascade of events the translocation of addi- tional glucose transporters (GLUT-4) to the cell membrane. New Engl J Med 341:248, 1999 GLUT-4 Translocation to the Plasma Membrane
Signal Transduction, Oncogenes and Cancer Tumor cells can express mutationally altered forms or levels of proteins involved in signal transduction – e.g. altered growth factors, growth factor receptors, G proteins, nuclear receptors, protein kinases, etc. Tumor cells may contain a normal signal transduction protein, but in excessive amounts. The responsible gene is called an Oncogene. Corresponding normal gene is a Protooncogene. How do protooncogenes become oncogenes? 1. Once a cellular “protooncogene” becomes part of a retroviral genome, it can undergo mutation, producing an oncogene, which can transform a normal cell subsequent to retroviral infection. 2. Spontaneous or chemically-induced (carcinogens) mutations in a “protooncogene.” 3. E.g., ras genes altered in codon 12,13 or 61 have been detected in 30% of spontaneous and chemic- ally induced tumors in animals and humans.
Most ras oncogene proteins lack GTPase activity. Several mutations that generate ras oncogenes are positioned close to the bound guanine nucleotide.
EGFR and Cancer • Several mechanisms by which the EGFR system • becomes oncogenic: • autocrine ligand loops • amplification or overexpression of the EGFR • receptor • deletions or mutations that render the receptor • constitutively active independent of ligands
Activities of EGFR are mediated by several signal transduction pathways. The best characterized is the Ras/Raf/Mek/Erk pathway, which leads to gene transcription resulting principally in cell growth and proliferation.
** Most common mutation produces EGFRvIII, which has a truncated extracellular region with a distorted ligand- binding area. Although this mutant cannot bind ligands, it is trapped in a partially activated state, unable to undergo downregulation. [(+) ligand dependence; (-) ligand independence]
EGFR as a Molecular Target for Cancer Therapy Extracellular MoAbs to EGFR MoAbs to EGFRvIII Anti-EGFRvIII vaccine Bispecific Abs Immunotoxin conjugates Ligand-toxin conjugates Intracellular Small molecular inhibitors of EGFR Tyr kinase Nuclear Antisense oligos Ribozymes
PTKRs and Bi-directional Signaling: Eph receptors and Ephrin ligands (Dodelet VC and Pasquale EB. Oncogene 19:5614-5619, 2000) Ephrin ligands are membrane-bound, with membrane association occuring through a GPI linkage (A) or as transmembrane proteins (B). Upon cell-cell contact, Eph receptors and ephrins engage in a class specific manner (A-A; B-B), which leads to receptor clustering, receptor activation and receptor phosphorylation (shown for EphB), which in turn provides docking sites for SH2 domain-containing signaling proteins. Upon receptor binding, ephrin-B ligands also become Phosphorylated on Tyr, via an unidentified associated tyrosine kinase. It is unknown if SH2 domain-containing proteins also dock to phosphorylated ephrin-B ligands. Ephrin/Eph interactions have been shown to be important in clot retraction mediated by activated platelets.
Tyrosine Kinase-associated Receptors Receptor Protein Tyrosine Kinases
Families of Ligands for Enzyme-linked Receptors Molecular Basis of Medical Cell Biology, 1998 Ligands for Tyrosine-kinase associated receptors
Ligands Activating Tyrosine Kinase-Associated Receptors Primarily involved in signaling between cells of the immune and hematopoietic systems, and signaling from cells near a site of inflammation
Tyrosine Kinase-Associated Receptors Redundant Signaling: Many members of the cytokine family can stimulate the same response in a cell Molecular basis of redundancy involves sharing of receptor transducing subunits Receptors composed of two polypeptide chains: Recognition subunit specific for and binds cytokine. Transducing subunit initiates signaling response.
The Interleukin-3 (IL-3) Receptor “Common” subunit transduces signal, which is shared by the IL-5 and GM-CSF receptors Subsequent to ligand binding, the receptor is phosphorylated on select Tyr residues by a special family of cytoplasmic Tyr kinases: Janus kinases (JAKs) of which there are four members (JAK1, JAK2, JAK3 and TYK2). These kinases appear to reside in close proximity to receptors on the inner surface of the membrane. These kinases also phosphorylate signaling molecules containing the required SH2/3 domains that dock on the receptor.
BASIC PRINCIPLES OF TYROSINE KINASE ASSOCIATED RECEPTOR SIGNALING I Subsequent to ligand binding, the receptor is phosphorylated on select Tyr residues by a special family of cytoplasmic Tyr kinases: Janus kinases (JAKs) of which there are four members (JAK1, JAK2, JAK3 and TYK2). These kinases appear to reside in close proximity to receptors on the inner surface of the membrane.