Download

1 / 41
410 likes | 427 Views
Explore the prediction of helical transmembrane proteins through statistical analysis and computational techniques. Understand the complex structure at residue and atomistic levels, crucial for drug development on membrane proteins.

E N D
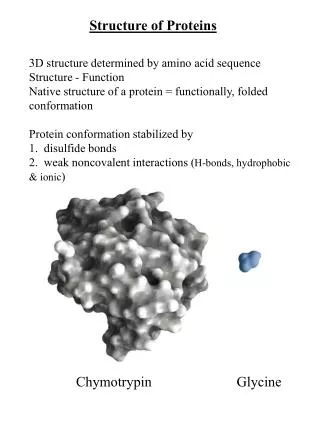
Multi-Scale Hierarchical Structure Prediction of Helical Transmembrane Proteins Zhong Chen and Ying Xu Department of Biochemistry and Molecular Biology and Institute of Bioinformatics University of Georgia
Outline • Background information • Statistical analysis of known membrane protein structures • Structure prediction at residual level • Helix packing at atomistic level • Linking predictions at residue and atomistic levels
Roles in biological process: Receptors; Channels, gates and pumps; Electric/chemical potential; Energy transduction > 50% new drug targets are membrane proteins (MP). Helical structure Beta structure Membrane Proteins
Membrane Proteins • 20-30% of the genes in a genome encode MPs. • < 1% of the structures in the Protein Data Bank (PDB) are MPs difficulties in experimental structure determination.
Membrane Proteins • Prediction for transmembrane (TM) segments (α-helix or β-sheet) based on sequence alone is very accurate (up to 95%); • Prediction of the tertiary structure of the TM segments: how do these α-helices/β-sheets arrange themselves in the constrains of bi-lipid layers? Helical structures are relatively easier to solve computationally
Membrane Protein Structures • Difficult to solve experimentally • Computational techniques could possibly play a significant role in solving MP structures, particularly helical structures
High Level Plan • Statistical analysis of known structures: • Unveil the underlying principles for MP structure and stability; • Develop knowledge-based propensity scale and energy functions. • Structure prediction at residue level • Structure prediction at atomistic level: MC, MD • multi-scale, hierarchical computational framework
Database for Known MP Structures: Helical Bundles • Redundant database • 50 pdb files • 135 protein chains • Non-redundant database (identity < 30%) • 39 pdb files • 95 protein chains (avg. length ~220 AA)
Bi-lipid Layer Chemistry Polar header (glycerol, phosphate) Hydrophobic tail (fatty acid)
Statistics-based energy functions • Length of bi-lipid layer: ~60Å • Central regions • Terminal regions • Three energy terms • Lipid-facing potential • Residue-depth potential • Inter-helical interaction potential Terminal 60 Å 30 Å Central Terminal
Lipid-facing Propensity Scale fraction of AAare lipid-facing LF_scale(AA) = fraction of AAare in interior • The most hydrophobic residues (ILE, VAL, LEU) prefer the surface of MPs in the central region, while prefer interior position in the terminal regions; • Small residues (GLY, ALA, CYS, THR) tend to be buried in the helix bundle; • Bulky residues (LYS, ARG, TRP, HIS) are likely to be found on the surface. This propensity scale reflects both hydrophobic interactions and helix packing
30 20 10 Y (Angstrom) 0 -10 -20 -30 -30 -20 -10 0 10 20 30 X (Angstrom) Helical Wheel and Moment Analysis The magnitude of each thin-vector is proportional to the LF-propensity and overall lipid-facing vector is the sum of all thin vectors, * Average Predication Error: 41 degree Lipid facing vector prediction: state of the art kPROT: avg. error ~41º Samatey Scale: 61º Hydrophobicity scales: 65 ~68º
Reside-Depth Potential - hydrophobic residues tend to be located in the hydrocarbon core; - hydrophilic residues tend to be closer to terminal regions; - aromatic residues prefer the interface region.
23º TM Helix Tilt Angle Prediction major pVIII coat protein of the filamentous fd bacteriophage (1MZT)
Statistical energy potentials (summary) • Three residue-based statistic potentials were derived from the database: (a) lipid-facing propensity, (b) residue depth potential, (c) inter-helical pair-wise potential • The lipid-facing scale predicted the lipid-facing direction for single helix with a uncertainty at ~ ±40º; • The residue-depth potential was able to predict the tilt angle for single helix with high accuracy. • Need more data to make inter-helical pair-wise potential more reliable
Key Prediction Steps • Structure prediction through optimizing our statistical potential (weighted sum) • Idealized and rigid helical backbone configurations; • Monte Carlo moves: translations, rotations, rotation by helix axis; • Wang-Landau sampling technique for MC simulation • Principle component analysis.
Wang-Landau Method for MC Observation: if a random walk is performed with probability proportional to reciprocal of density of states then a flat energy histogram could be obtained. The density of states is not known a priori. In Wang-Landau, g(E) is initially set to 1 and modified “on the fly”. Monte Carlo moves are accepted with probability Each time when an energy level E is visited, its density of states is updated by a modification factor f >1, i.e.,
Wang-Landau Method for MC • Advantages: • simple formulation and general applicability; • Entropy and free energy information derivable from g(E); • Each energy state is visited with equal probability, so energy barriers are overcome with relative ease.
Principal Component Analysis • Purpose: • analyze the conformation variations during a simulation, and • identify the most important conformational degrees of freedom. • Covariance matrix: * A large part of the system’s fluctuations can be described in terms of only a few PCA eigenvectors.
A Model System: Glycophorin (GpA) Dimer • GxxxG motif • Ridges-into-grooves 22 residues, 189 atoms EITLIIFGVMAGVMAGVIGTILLISY
Glycophorin (GpA) Dimer (1AFO) A: GEM (global energy minimum) RMSD=3.6A E=-114.6kcal/mol B: LEM RMSD=0.8A E=-93.9kcal/mol B A RED: experiment GREY: simulation
Helices A and B of Bacteriorhodopsin (1QHJ) A B A: GEM RMSD=2.7A E=-94kcal/mol B: LEM RMSD=0.9A E=-86kcal/mol RED: experiment GREY: simulation
Bacteriorhodopsin (1QHJ) Rmsd=5.0A G F A A E C B D Computational prediction Experimental structure
Residue-level structure prediction (Summary) • A computational scheme was established for TM helix structure prediction at residue level; • For two-helix systems, LEM structures very close to native structures (RMSD < 1.0 Å) were consistently predicted; • For a seven-helix bundle, a packing topology within 5.0 Å of the crystal structure was identified as one of the LEMs.
Key Prediction Steps • Structure prediction through optimizing atom-level energy potential: • CHARMM19 force field for helix-helix interaction • Knowledge-based energy function for lipid-helix interaction • Idealized and rigid helix structure for backbone andsidechain flexible; • Apply helix orientation constraint (i.e., N-term inside/outside cell); • MC moves: translations, rotations, rotation by helix axis, and side-chain torsional rotation; • Wang-Landau algorithm for MC simulation
CHARMM19 Polar Hydrogen Force Field - nonpolar hydrogen atoms are combined with heavy atoms they are bound to , - polar hydrogen atoms are modeled explicitly.
Effect of Helix-Lipid Interactions: Helices A&B of Bacteriorhodopsin Helix-helix interactions Helix-helix & helix-lipid interactions Helix-lipid interactions play a critical role in the correct packing of helices
RMSD=0.2Å RMSD=4.4Å RMSD=5.7Å RMSD=7.1Å Effect of Helix-Lipid Interactions: Helix A&B of Bacteriorhodopsin (BR) Hydrocarbon core region 30 Å All four LEM structures share essentially the same contact surfaces. In the native structure, the polar N-terminals of both helices are located outside of hydrocarbon core region, resulting in low helix-lipid energy.
Docking of a Seven-helix Bundle: Bacteriorhodopsin (1QHJ) Crystal structure 7 helices, 174 residues, 1619 atoms A • CHARMM19 + lipid-helix potential; • One month CPU time on one PC B B A Initial Configuration
Rmsd=4.7A Rmsd=3.0A Rmsd=6.6A Rmsd=8.0A Rmsd=8.4A Potential Energy Landscape
Global Energy Minimum Structure (RMSD=3.0 Å) RED: experiment GREY: simulation
Atom-level Structure Prediction (Summary) • Wang-Landau algorithm proved to be effective for the energetics study of TM helix packing; • Prediction results for two-helix and seven-helix structures are highly promising • Practical application of Wang-landau method to large systems requires further work.
Part IV: Linking Predictions at Residue- and Atomistic levels
Correspondence between simulations at two levels • A multi-scale hierarchical modeling approach is feasible and practical: • LEMs identified at residue-level be used as candidates for atomistic simulation; • Using PC vectors from residue-level simulation to improve search speed in atomistic simulation.
Future Works • Further improvement of the residue-based folding potentials; • Speed-up and parallelization of Wang-Landau sampling; • Construct a hierarchical computational framework, and develop corresponding software package.
Acknowledgements • Funding from NSF/DBI, NSF/ITR, NIH, and Georgia Cancer Coalition • Dr. David Landau (Wang-Landau algorithm) and Dr. Jim Prestegard (NMR data generation) of UGA • Thanks DIMACS for invitation to speak here