Download
1 / 56
560 likes | 746 Views
专业词汇. pharmacist n. 配药者 , 药剂师 elucidate vt. 阐明 , 说明 curare n. 箭毒马鞍子 , 箭毒 ( 箭毒马鞍子的毒素 , 南美印地安人用以浸制毒箭 ) toxic adj. 有毒的 , 中毒的 antagonistic adj. 颉颃的 vagus n. 迷走神经 Vagusstoff n. 迷走神经释放物. acetylcholine (ACh) n. 乙酰胆碱 ray n. 鳐 potential prey 潜在的食物 acetylcholinesterase n. 乙酰胆碱酯酶

E N D
专业词汇 • pharmacist n.配药者, 药剂师 • elucidate vt. 阐明, 说明 • curare n.箭毒马鞍子,箭毒(箭毒马鞍子的毒素,南美印地安人用以浸制毒箭) • toxic adj.有毒的, 中毒的 • antagonistic adj.颉颃的 • vagus n.迷走神经 • Vagusstoff n.迷走神经释放物
acetylcholine (ACh) n.乙酰胆碱 • ray n.鳐 • potential prey 潜在的食物 • acetylcholinesterase n.乙酰胆碱酯酶 • motor nerves n.运动神经 • acetylcholine receptor (nAChR) n.乙酰胆碱受体 • parasympathetic nerve 副交感神经 • synapses n.突触 • atropine n.阿托品
postsynaptic neurons n.突触后神经元 • bungarotoxin n.金环蛇毒素 • venom n.蛇毒 • homogenize vt.使均质 • blender n. 搅拌机 • centrifuging n.离心过滤 • pellet n. 小球 • nonionic detergent非离子型洗涤剂 • hydrophobic adj.疏水性的 • hydrophilic adj.亲水的, 吸水的
phospholipid n.磷脂 • disruptive adj.使破裂的, 分裂性的 • carboxyl n.羧基 • resemblance n.相似之处 • affinity chromatography n.亲合色谱法(层析) • pentamer n.五聚物 • electrophoresis n.电泳, 现象 • polyacrylamide gel n.聚丙烯酰胺凝胶
cation n.阳离子 • postulate n.假定 • conformational n.构象的 • inward flux 内流 • depolarization n.去极化 • sodium n.钠 • potassium n.钾 • mutagenesis n.诱变 • neurotransmitter n.神经递质
ethane n.乙烷 • spray vt.喷射 • grid n. 格子, 栅格 • peripheral adj.外围的 • agrin n.集聚蛋白,集聚素[由运动神经分泌并可诱 导肌纤维的乙酰胆碱酯酶和乙酰胆碱受体发生聚集] • axon n.轴突 • a cascade of events n.级联放大反应
in the vicinity of adv.在邻近 • phosphorylation n.磷酸化 • kinase n.激酶 • polymerization n.聚合 • sepharose beads n.琼脂糖凝胶珠 • affinity column n.亲和层析柱 • flaxedil n.三乙碘化三苯 • elute vt.洗脱
发现历程 • 19c Claude Bernard:实验毒物及麻醉学说 • 1906 John Langley:化学递质假说 • 1921 Loewi:发现化学递质(ACh) • 1937 David Nachmansohn:发现nAChR的超级储存库(EO) • 19c70y 分离纯化nAChR • 19cL70y nAChR结构的揭示 • nAChR的精细结构及其作用机理 • 1980s U.J. McMahan:agrin诱导聚集nAChR的机制
19c Claude Bernard:实验毒物及麻醉学说 • 伯纳尔Bernard,Claude (1813—1878)。法国生理学家,生于隆恩(Rhǒne)圣瑞立安(St.Julien)。在1843年,30岁的Claude Bernard已是一名药剂师并兼职一份很有前途的剧作家工作,但为了成为一名文学家的梦想,他离开了家乡,来到巴黎。 • 但是,到巴黎后,一个偶然的机会使Bernard继续进修医学,并任法国大学(Collége deFrance)生理学教授马迪让(F.Magendie)的助手,开始了研究生活。他的研究涉及到生理学的广泛领域,被后人认为是实验医学、普通生理学的创始人。
curare • curare是一种取自热带植物的 剧毒药物,被南美的土著猎人用 作制作毒箭的材料,因为它能麻 痹肌肉。 • Bernard发现 curare在麻痹骨胳肌的过程中,既不影响神经传导冲动到肌肉的能力,也不影响肌肉受到刺激收缩的能力。于是,Bernard认为curare 的作用位点是神经与肌肉相接触的区域(NMJ)。 • 这一结论后来被John Langley 证实并进一步补充和完善。
1906 John Langley:化学递质假说 • Langley 当时正在研究另一种植物提取物烟碱(nicotine)的特性。他发现nicotine 能刺激蛙的离体骨骼肌的收缩,但curare 会使nicotine 失效。 • 在1906年,Langley提出:冲动从神经传递到肌肉,并不是借助于物理方法,就像电流在两根电线之间流过一样,而是一种特殊物质从神经末稍释放的结果。 • 他把这一传递物称为化学递质,并认为化学递质通过与肌细胞表面的受体物质结合而传导信号,就是在此位点结合了nicotine与curare。这些论断在后来被证明是很有远见的。
Langley的刺激通过神经释放化学物质传递到肌肉的假说在1921年被奥地利生理学家Otto Loewi的一个设计精巧的实验证明。
1921 Loewi:发现化学递质(ACh) • 化学家凯库勒(Friedrich August Kekulé)在梦中悟出苯环的结构,是大众熟知的故事 。
灵感之梦 • 1864年的某个冬天,德国化学家凯库勒坐在壁炉前打了个瞌睡,原子和分子们开始在幻觉中跳舞,一条碳原子链像蛇一样咬住自己的尾巴,在他眼前旋转。猛然惊醒之后,凯库勒明白了苯分子是一个环——就是现在充满了我们的有机化学教科书的那个六角形的圈圈。 • 这个最著名的例子并不是唯一。 • Loewi的精巧实验也源于梦。
1921年复活节星期天之前的那个夜晚,奥地利生物学家洛伊(Otto Loewi)从梦中醒来,打开灯,抓过一张纸迷迷糊糊地写了些东西,倒下去又睡着了。早上6点钟,他突然想到,自己昨夜记下了一些极其重要的东西,赶紧把那张纸拿来看,却怎么也看不明白自己写的是些什么鬼画符。幸运的是,第二天凌晨3点,逃走的新思想又回来了。
它是一个实验的设计方法,可以用来验证洛伊17年前提出的某个假说是否正确。它是一个实验的设计方法,可以用来验证洛伊17年前提出的某个假说是否正确。 • 洛伊赶紧起床,跑到实验室,杀掉了两只不幸的青蛙,取出蛙心泡在生理盐水里,其中一号带着迷走神经,二号不带。 用电极刺激第一号 心脏的迷走神经使 心脏跳动变慢,几 分钟后把泡着它的 盐水移到二号心脏 所在的容器里,结 果二号心脏的跳动也放慢了。
这个实验表明,神经并不直接作用 于肌肉,而是通过释放化学物质来 起作用,一号心脏的迷走神经受刺 激时产生了某些物质,它们溶解在 盐水里,对二号心脏产生了作用。 Loewi把这种能抑制蛙的心脏跳动的物质称作“Vagusstoff” 。 • 经过几年的研究,Loewi发现这种物质的化学及生理学性质与乙酰胆碱完全相同,于是他断定是迷走神经末稍释放的是乙酰胆碱(ACh) 。
神经冲动的化学传递就这样被发现了,它开启了一个全新的研究领域,并使洛伊获得1936年诺贝尔生理学或医学奖。神经冲动的化学传递就这样被发现了,它开启了一个全新的研究领域,并使洛伊获得1936年诺贝尔生理学或医学奖。
1937 David Nachmansohn:发现nAChR的超级储存库(EO) • 1937年,正当梭尔邦(Sorbonne)大学的神经生理学家David Nachmansohn参观巴黎世界博览会时,他注意到有几只具发电器官(electric organ, EO)的鳐正在表演节目。这些鳐的EO能够发出40~60V的电压,杀死水中的潜在食物。
OE • 发电器包括大量的电板 (最高可达20 0万块), 每一块电板来自一个特化 的多核肌细胞,电板之间 隔以胶状物质,许多电板 包在一个结缔组织构成的小室内,神经通过结缔组织分支到每一电板。电鳐的电板呈垂直排列,放电的方向是由腹侧向背侧;电鳗的电板呈水平排列,放电的方向是由尾部向头部。
当时Nachmansohn正在研究乙酰胆碱酯酶(AChase),AChase可酶解从运动神经末稍释放的ACh。当时Nachmansohn正在研究乙酰胆碱酯酶(AChase),AChase可酶解从运动神经末稍释放的ACh。 • Nachmansohn知道这类鱼的EO与骨骼肌是同源的,于是在博览会结束后,开始对EO进行研究。 • 对EO的第一次实验结果表明它是AChase的超级储存库。此器官也是nAChR十分丰富的储存库,nAChR存在于骨骼肌细胞的突触后膜上,它会与由运动神经末稍释放的ACh分子结合。
如果能发现一个理想的系统模型,对于细胞结构和功能的特殊领域的研究,可以说是无价的。这将在后面的讨论中得以证实,鱼的发电器官事实上是nAChR研究中的唯一物质来源。如果能发现一个理想的系统模型,对于细胞结构和功能的特殊领域的研究,可以说是无价的。这将在后面的讨论中得以证实,鱼的发电器官事实上是nAChR研究中的唯一物质来源。
19c70y 分离纯化nAChR • Chen-Yan Lee:发现nAChR的理想标记物 • nAChR的分离纯化
α-BGT:nAChR的理想标记物 • 由于nAChR是一种整合膜蛋白,直到19世纪70年代,才发展起了分离此类蛋白的技术。 • 一种特殊蛋白的提纯需要一种与之亲和的标记物,来检测其在所分离的蛋白质片断中的纯度。 • 作为nAChR的理想标记物的是能选择性的与其紧密结合的一种化合物。这种化合物被台湾国立大学的Chen-Yuan Lee和他的同事们在1963年发现,是从台湾本土的一种毒蛇的毒液中提取出的,叫做α-银环蛇毒素(α-BGT)。α-BGT通过与骨骼肌细胞突触后膜的nAChRs紧密结合,而阻碍ACh的传递,造成肌肉麻痹。
nAChR的分离纯化 • 在人们有α-BGT作为nAChR的标记物,EO作原料,以及一种溶解膜蛋白的去污剂之后,nAChR才得以在19世纪70年代分离。 • 在分离纯化的研究中,有一种方法是这样的:
①得到含nAChR的细胞膜 • 先用搅拌器将EO的一部分制备成均一化的悬浮液,然后离心,使悬浮液中的质膜片断形成颗粒状物。
②提取膜蛋白 • 经过许多溶解膜蛋白的尝试,最后发现TritonX-100的效果最佳。 • TritonX-100是一种非离子型去污剂。它与其他去污剂的相同点是都具有一个长的能取代膜蛋白上磷脂的疏水基团,以及一个能够溶解于水的亲水末端;不同的是大多数其它去污剂都带有一个带负电的羟基基团,而它没有,所以是非离子型的,对蛋白质结构的破坏性小。 • 于是,膜蛋白就可以用TritonX-100从质膜片断中提取出来。
③分离纯化nAChR • 将膜蛋白混合物灌注到装有人造复合物—琼脂糖珠的层析柱中,这些琼脂糖珠的末端都带有一个ACh的结构类似物作为表面吸附剂。(Figure2a) • 当这些膜蛋白流经层析柱时,两种与ACh有亲和位点的蛋白,nAChR和AChE,他们就被吸附在琼脂糖珠上。剩下约90﹪的其它膜蛋白由于不能被吸附,直接流经层析柱。(Figure2b)
再用10-3M的洗脱液flaxedil(三乙碘化三苯(β-二乙氨乙氧基))洗脱,nAChR会被选择性地洗脱下来,并被收集,最后剩下AChE。采用这一方法后,用α-BGT做标记物的检测结果表明,AChRs在一步中被纯化了150多倍。这种方法就是亲和层析。再用10-3M的洗脱液flaxedil(三乙碘化三苯(β-二乙氨乙氧基))洗脱,nAChR会被选择性地洗脱下来,并被收集,最后剩下AChE。采用这一方法后,用α-BGT做标记物的检测结果表明,AChRs在一步中被纯化了150多倍。这种方法就是亲和层析。
19cL70y nAChR结构的揭示 • 哥伦比亚大学的Arthur Karlin经过实验研究发现nAChR是一个五聚体,即一个由5个亚基组成的蛋白质。每一个受体都包含有2个α亚基,β、γ、δ亚基各一个。亚基的分离提纯是在前面步骤的基础上,用聚丙烯酰胺凝胶电泳的技术方法完成,主要是按各多肽(亚基)的大小将其分离。(Figure3)
对nAChR的另一个具有里程碑意义的研究结果是证明了被提纯的受体蛋白除了有一个与ACh相结合的位点外,还具有一个阳离子通道。对nAChR的另一个具有里程碑意义的研究结果是证明了被提纯的受体蛋白除了有一个与ACh相结合的位点外,还具有一个阳离子通道。 • 这个结论的假说已在此几年前,由巴黎巴斯德研究所的Jean-Pierre Changeux提出:当ACh与nAChR结合后,会使nAChR的构象发生改变,使离子通道开放。这样,由于细胞膜内外Na+离子的浓度差,导致Na+内流,膜去极化,骨骼肌收缩。
在19世纪70年代后半期,Changeux和他的同事们成功地将纯化的nAChR分子加到脂质体(人造磷脂泡)上。他们用做好标记的含有不同Na+、K+浓度的脂质体证明:ACh与nAChR在磷脂双分子成的结合会使膜内外的阳离子发生流动。在19世纪70年代后半期,Changeux和他的同事们成功地将纯化的nAChR分子加到脂质体(人造磷脂泡)上。他们用做好标记的含有不同Na+、K+浓度的脂质体证明:ACh与nAChR在磷脂双分子成的结合会使膜内外的阳离子发生流动。 • 很显然,这种受体蛋白确实具有传递化学信号的所有结构部件:一个与ACh的结合位点;一个门控离子通道,以及二者之间的协调机制。
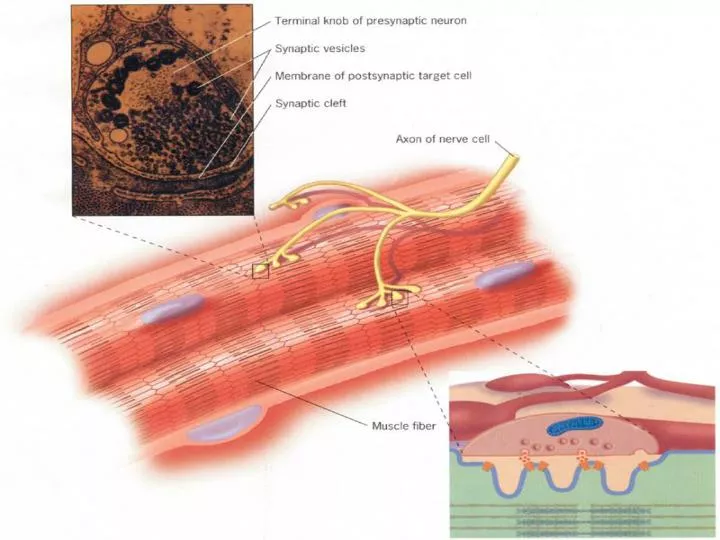
nAChR的精细结构及其作用机理 • 对于结构的分析,有许多不同的途径。其中的一条是采用基因纯化,氨基酸测序和定点诱变的方法来确定多肽的跨膜位点,神经递质的结合位点以及形成离子通道的位点。
另一种途径则需借助电子显微镜。第一次瞥见nAChR是在电镜下观察发电器官细胞的质膜时(Figure4)。另一种途径则需借助电子显微镜。第一次瞥见nAChR是在电镜下观察发电器官细胞的质膜时(Figure4)。 • nAChR看起来是环形的,具有约8nm的直径和一个2nm直径的中央通道,并且它从磷脂双分子层突出到细胞外空间。
nAChR更精确的模型已经被英格兰医学研究所的Nigel Unwin和他的同事们所描绘。经过对EO冷冻膜电镜照片的数学分析,Unwin将nAChR的组成描述为5个亚基围绕着一个中央通道。(Figure5)
离子通道是一个很窄的直径只有 • 7~8Å的孔道,孔道被5个亚基 • 的α-helix片断组成的墙所包围。 • 孔道的门控开关被认为是在跨膜 • 通道的中央附近,在这里,5个 • α-helix屈向中心,形成一个结。 • 在这个模型中, • 5个α-helix的 • Leu(亮氨酸) 残基组成了一个紧缩环,限制 离子的跨膜流动。当ACh分子 与两个α亚基膜外结构域口袋 结构的一个位点结合后,门控 开关被打开。
为了研究通道开放过程中nAChR的变化,Unwin做了下面的实验。为了研究通道开放过程中nAChR的变化,Unwin做了下面的实验。 • 将准备好的富含nAChR的细胞膜涂抹在一个支持网络上,并使它能够流到装有能使细胞膜冻结的液氮乙烷的容器中。在nAChR溶液到达冷冻池前约5毫秒(msec)时,向网格喷射ACh溶液,ACh就与受体结合并导致其构象变化而打开通道。
通过比较nAChR通道开放与关闭的电镜照片,Unwin发现ACh的结合触发了受体亚基细胞膜外结构域的构象发生了改变,并且传播到整个受体蛋白,导致围绕在孔道周围的α-helix构象改变。(Figure6)孔道的封闭式由于α-helix处结的形成,这一点可从Figure6b的白、蓝条比较看出。通过比较nAChR通道开放与关闭的电镜照片,Unwin发现ACh的结合触发了受体亚基细胞膜外结构域的构象发生了改变,并且传播到整个受体蛋白,导致围绕在孔道周围的α-helix构象改变。(Figure6)孔道的封闭式由于α-helix处结的形成,这一点可从Figure6b的白、蓝条比较看出。
疑问 • 我们知道膜外周的部件可限制膜整合蛋白的移动。在nAChR上也发现了一个典型的例子。在胚胎早期,肌肉无神经支配时,nAChR均匀分布在肌肉表面。而成熟神经肌肉接头处nAChR则高密度聚集。 • 运动神经末梢下的突触区小于肌肉细胞表面积的0.1﹪,却集中了约90﹪的nAChR。这是为什么呢?
1980s U.J. McMahan:agrin诱导聚集nAChR的机制 • 在19世纪80年代,斯坦福大学的U.J. McMahan和他的同事们在EO中发现了一种蛋白,当把这种蛋白加入培养的未成熟肌细胞中后,它能诱导nAChRs在其细胞膜的聚集。并且还发现它在EO的突触间隙处有很高的浓度。 • 1987年,他们纯化了这种蛋白,并将它命名为集聚蛋白(agrin)。
McMahan提出agrin是由运动神经合成,延轴突运输到终端神经节,并在此被释放到神经肌肉接头处的细胞外空间。这些agrin分子而后与邻近肌细胞质膜上的特异性agrin受体结合,触发一系列级联放大反应,使nAChR分子(以及突触后膜的其他组成部分)聚集到活化的agrin受体附近,并达到最大浓度。McMahan提出agrin是由运动神经合成,延轴突运输到终端神经节,并在此被释放到神经肌肉接头处的细胞外空间。这些agrin分子而后与邻近肌细胞质膜上的特异性agrin受体结合,触发一系列级联放大反应,使nAChR分子(以及突触后膜的其他组成部分)聚集到活化的agrin受体附近,并达到最大浓度。 • Agrin作为突触间隙细胞外物质的一部分,帮助维持后突触膜的结构,并保证新合成的nAChRs在突触位点的富集。
多种研究都支持这一假说,包括利用抗体与agrin结合的特异性所进行的研究。多种研究都支持这一假说,包括利用抗体与agrin结合的特异性所进行的研究。 • 如果使鸡胚胎脊髓的一段,携带抗agrin的抗体,此抗体就会特异性地标记运动神经的细胞质,这表明agrin分子来源于此。抗体当然也存在于神经肌肉接头处的突触间隙。若将此抗体加入到鸡的运动神经和正在生长的肌细胞的混合培养物中,所培养的两类细胞接触位点处nAChR的正常聚集将会被阻断。
更深入的关于神经肌肉接头(NMJ)形成的问题,由一项对鼠的agrin基因敲除实验解决。即通过遗传工程的方法使鼠的运动神经不能产生agrin。Agrin缺陷鼠在母体中发育正常,但在妊娠期却死亡。更深入的关于神经肌肉接头(NMJ)形成的问题,由一项对鼠的agrin基因敲除实验解决。即通过遗传工程的方法使鼠的运动神经不能产生agrin。Agrin缺陷鼠在母体中发育正常,但在妊娠期却死亡。 • 这些鼠在死前没有任何肌肉运动的迹象出现,并且在肌肉组织的检查中,发现缺少正常的神经肌肉突触。
Agrin诱导聚集nAChR的机制,对我们来说,仍然知之甚少,但这一过程中的许多步骤已被界定。将agrin加入到未成熟的肌细胞后,发生的事件之一是nAChR β亚基的一个或多个酪氨酸(Tyr)残基的磷酸化。 • 这个磷酸化作用发生在出现nAChR聚集的迹象之前很长一段时间。 • Tyr的磷酸化在细胞膜信号传导的过程中常是一个关键步骤。如果nAChR的Tyr磷酸化被加入的Tyr激酶失活物阻断,nAChR将不再聚集,这表明agrin诱导nAChR的Tyr磷酸化在聚集作用中是一个不可缺少的步骤。
信号的传导路径由于Tyr的磷酸化而被激活,并常会导致细胞骨架元件的重排,特别是肌动蛋白纤维。细胞骨架经常是连在肌动蛋白骨架(AS)上的,AS在整合膜蛋白的分配上扮演着一个关键角色。信号的传导路径由于Tyr的磷酸化而被激活,并常会导致细胞骨架元件的重排,特别是肌动蛋白纤维。细胞骨架经常是连在肌动蛋白骨架(AS)上的,AS在整合膜蛋白的分配上扮演着一个关键角色。 • 最近的研究显示,细胞表面信号的产生是在agrin和它的受体结合导致AF去极化之后,这被认为是nAChR聚集的直接原因。如果AF的形成被能与actin单体结合并阻止单体间联合形成多聚体的药物阻断,agrin诱导nAChRs的聚集就会被阻止。
目前公认的导致AChR在突触后膜高密度聚集途径目前公认的导致AChR在突触后膜高密度聚集途径 • Agrin/ MuSK信号转导通路 • Neuregulin/ErbB信号转导通路
Agrin/ MuSK信号转导通路 • 集聚蛋白(Agrin)主要由运动神经元合成,经神经末梢释放并沉积于基底层,触发AChR聚集,而不增加AChR的数量。 • 肌肉的受体酪氨酸激酶(muscle-specific receptor tyrosine kinase, MuSK)是Agrin的受体,选择性在骨骼肌表达,与AChR共同聚集于成年神经肌肉接头的突触后膜。 • 受Agrin的影响,MuSK被快速磷酸化,介导神经肌肉接头nAChR的磷酸化。
此过程还必须依赖于突触后膜ACh受体缔合蛋白(acetylcholine receptor-associated protein at synapse,Rapsyn)的存在。 Rapsyn主要在肌肉组织合成,是一种细胞骨架蛋白。它在体内与神经肌肉接头nAChR以等摩尔数存在,共同定位 。 • 上述3种分子密切相连,神经轴突分泌Agrin后,在Rapsyn的参与下MuSK被磷酸化,使nAChR磷酸化,由此诱导AChR在终板膜聚集。
