Download
1 / 1
10 likes | 143 Views
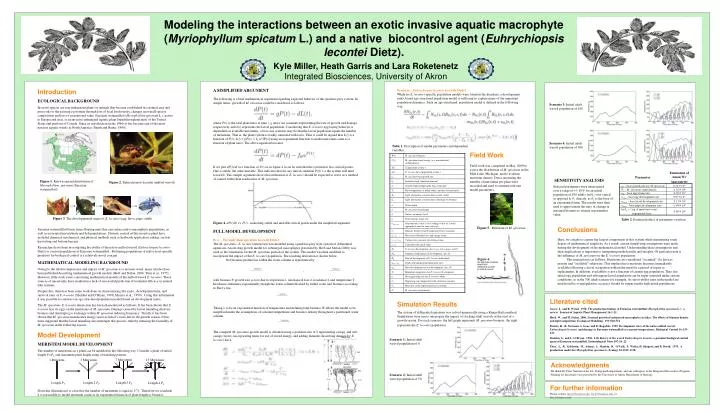
Modeling the interactions between an exotic invasive aquatic macrophyte ( Myriophyllum spicatum L.) and a native biocontrol agent ( Euhrychiopsis lecontei Dietz). Kyle Miller, Heath Garris and Lara Roketenetz Integrated Biosciences, University of Akron. Introduction ECOLOGICAL BACKGROUND

E N D
Modeling the interactions between an exotic invasive aquatic macrophyte (Myriophyllumspicatum L.) and a native biocontrol agent (Euhrychiopsislecontei Dietz). Kyle Miller, Heath Garris and Lara RoketenetzIntegrated Biosciences, University of Akron Introduction ECOLOGICAL BACKGROUND Invasive species are non-indigenous plants or animals that become established in a natural area and pose risks to the existing ecosystem through loss of local biodiversity, changes in overall species composition and loss of recreational value. Eurasian watermilfoil (Myriophyllumspicatum L.), native to Europe and Asia, is an invasive submerged aquatic plant found throughout most of the United States and portions of Canada. Since its introduction in the 1940s it has become one of the most noxious aquatic weeds in North America (Smith and Barko, 1990). Eurasian watermilfoil forms dense floating mats that can reduce native macrophyte populations, as well as associated invertebrate and fish populations. Historic control of this invasive plant have included chemical, mechanical, and physical methods such as herbicide application, raking, suction harvesting and bottom barriers. Researchers have been investigating the ability of the native milfoil weevil (Euhrychiopsislecontei Dietz) to control populations of Eurasian watermilfoil.. Bolstering populations of native host-specific predators for biological control is a relatively novel concept. MATHEMATICAL MODELING BACKROUND Owing to the relative importance and impact of M. spicatum as a noxious weed, many articles have been published describing mathematical growth models (Herb and Stefan, 2006; Titus et al., 1975). However, little work exists concerning mathematical models of the milfoil weevil E. lecontei. These sources of uncertainty have resulted in a lack of successful prediction of treatment efficacy in natural lake systems. Despite this, there has been some work done on characterizing life cycle, development time, and survival rates of E. lecontei (Sheldon and O’Bryan, 1996; Mazzei et al., 1999). Using this information it was possible to construct an age-structured population model based on development index. The M. spicatum–E. lecontei interaction has been characterized as follows. It has been shown that E. lecontei lays its eggs on the meristems of M. spicatum. Damage caused by larval tunneling destroys biomass and interrupts gas exchange within M. spicatum reducing buoyancy. Finally, it has been shown that M. spicatum translocates energy reserves into it’s roots late in the growth season. It has been suggested that the larval tunneling also interrupts this process, thereby reducing the fecundity of M. spicatum in the following season. Model Development MERISTEM MODEL DEVELOPMENT The number of meristems on a plant can be modeled in the following way. Consider a plant of initial length P=P0 and increment plant length using a branching pattern. From this illustration it is clear that the number of meristems is equal to 2P-1. Therefore we conclude it is reasonable to model meristem count as an exponential function of plant length or biomass. A SIMPLIFIED ARGUMENT The following is a brief mathematical argument regarding expected behavior of this predator prey system. In simple terms, growth of M. spicatum could be considered as follows. where P(t) is the total plant mass at time t, g and d are constants representing the rate of growth and damage respectively, and L(t) represents the larval population. Considering that E. lecontei egg laying behavior is dependent on available meristems, a best case scenario may be that the larval population equals the number of meristems. That is, the plant system is totally saturated with larva. Thus it could be argued that L(t) is a function of P(t); L(t) = f(P(t)) = f0^[P(t)] using an exponential function to model meristem count as a function of plant mass. The above equation becomes If we plot dP(t)/dt as a function of P(t) as in figure 4 it can be seen that the system has two critical points. One is stable, the other unstable. This indicates that for any initial condition P(0) > a the system will tend toward b. This simple argument shows that utilization of E. lecontei should be expected to serve as a method of control rather than eradication of M. spicatum. Figure 4.dP(t)/dt vs. P(t) - indicating stable and unstable critical points under the simplified argument. FULL MODEL DEVELOPMENT Prey - Myriophyllum spicatum Growth Model The M. spicatum - E. lecontei interaction was modeled using a predator-prey style system of differential equations. An existing growth model for submerged macrophytes presented by Herb and Stefan (2006) was used as the foundation for the M. spicatum portion of the system. This model was then modified to incorporate the impact of the E. lecontei population. The resulting derivation is shown below. Net biomass production within the water column is represented by with biomass P, growth rate , loss due to respiration , mechanical loss , irradiance I, and temperature T. Irradiance attenuates exponentially though the water column blocked by turbid water and biomass according to Beer’s law. Taking to be an exponential function of temperature and defining total biomass W allows the model to be simplified under the assumptions of constant temperature and biomass density throughout a partitioned water column. The complete M. spicatum growth model is obtained using a partition size of 2 representing canopy and sub-canopy layers, incorporating terms for use of stored energy, and adding elements describing damage by E. lecontei larva. Predator - Euhrychiopsis lecontei Growth Model While no E. lecontei-specific population models were found in the literature, a development index based age-structured population model is sufficient to capture many of the important population dynamics. Such an age-structured population model is defined in the following way. Simulation Results The system of differential equations was solved numerically using a Runge Kutta method. Simulations were run to interrogate the impact of stocking adult weevils at the start of a growth season. For each scenario, the left graph represents M. spicatum biomass, the right represents the E. lecontei population. Conclusions Here, we sought to capture the largest components of this system while maintaining some degree of mathematical simplicity. As a result, certain simplifying assumptions were made during the development of the mathematical model. Understanding these assumptions and their implications is important to interpreting model results and insights. Of particular note is the influence of M. spicatum on the E. lecontei population. This interaction is as follows. Meristems are considered “occupied” if a larva is present and “available” otherwise. This implies that a meristem becomes immediately available following a larval occupation without the need for a period of repair or replacement. In addition, availability is not a function of current egg population. Thus, the initial egg population and subsequent larval population can be super saturated under certain conditions, as in the 300 adult scenario for example. As survivability rates in the model are unaffected by overpopulation, accuracy should be suspect under high initial populations. Scenario 3: Initial adult weevil population of 100 Scenario 4: Initial adult weevil population of 300 Table 1. Description of model parameters and dependent variables. Field Work Field work was completed in May, 2009 to assess the distribution of M. spicatum in Six Mile Lake, Michigan, and to evaluate meristem density. Data concerning the number of meristems per plant were recorded and used to estimate relevant model parameters. SENSITIVITY ANALYSIS Figure 1. Known current distribution of Myriophyllumspicatum (Eurasian watermilfoil) Figure 2. Euhrychiopsis lecontei (milfoil weevil) Selected parameters were interrogated over a range of +/- 10% for an initial population of 100 adults. ln(θw) was varied as opposed to θw directly, as θw is the base of an exponential term. The results were then used to approximate the ratio of change in maximal biomass to change in parameter value. Figure 3. The developmental stages of E. lecontei (egg, larva, pupa, adult) Table 2. Estimated effect of parameter variation. Figure 5. Meristem of M. spicatum Figure 6. , Literature cited Grace, J., and R. Wetzel. 1978. The production biology of Eurasian watermilfoil (Myriophyllumspicatum L.): a review. Journal of Aquatic Plant Management 16:1–11. Herb, W., and H. Stefan. 2006. Seasonal growth of submersed macrophytes in lakes: The effects of biomass density and light competition. Ecological Modelling . 193:560-574 Mazzei, K., R. Newman, A. Loos, and D. Ragsdale. 1999. Development rates of the native milfoil weevil, Euhrychiopsislecontei, and damage to Eurasian watermilfoil at constant temperatures. Biological Control 16:139–143. Sheldon, S., and L. O’Bryan. 1996. Life history of the weevil Euhrychiopsislecontei, a potential biological control agent of Eurasian watermilfoil. Entomological News 107:16–22. Titus, J., R. Goldstein, M. Adams, J. Mankin, R. O’Neill, P. Weiler, H. Shugart, and R. Booth. 1975. A production model for Myriophyllumspicatum L. Ecology 56:1129–1138. Scenario 1: Initial adult weevil population of 0 7 Meristems 1 Meristem 15 Meristems 3 Meristems Acknowledgments We thank Dr. Peter Niewiarowski, Dr. Young math department,and our colleagues in the Integrated Biosciences Program. Funding for this project was provided by the University of Akron Department of Biology. Scenario 2: Initial adult weevil population of 50 Length 3 P0 Length P0 Length 2 P0 Length 4 P0 For further information Please contact jkm29@uakron.edu, hwg3@uakron.edu, or ldr11@uakron.edu