Download

1 / 1
10 likes | 110 Views
LIN12/NOTCH REPEATS (LNRs): IMPARTING TUNABLE REDOX RESPONSIVENESS IN LESS THAN 35 AMINO ACIDS THROUGH AN INTERPLAY BETWEEN Ca 2+ BINDING AND DISULFIDE BOND FORMATION Janelle L. Jackson, Angie Seo, Didem Vardar Ulu, Wellesley College, Chemistry Department, Wellesley, MA, USA. Introduction:

E N D
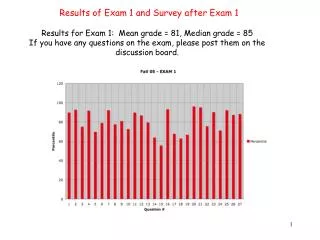
LIN12/NOTCH REPEATS (LNRs): IMPARTING TUNABLE REDOX RESPONSIVENESS IN LESS THAN 35 AMINO ACIDS THROUGH AN INTERPLAY BETWEEN Ca2+ BINDING AND DISULFIDE BOND FORMATION Janelle L. Jackson, Angie Seo, Didem Vardar Ulu, Wellesley College, Chemistry Department, Wellesley, MA, USA. Introduction: Many multi-domain proteins contain small protein modules whose global folds are stabilized by metal binding or disulfide bonds rather than an extensive hydrophobic core or secondary structures. Lin-12/Notch Repeat (LNR) is such a module, first identified in Notch receptors and more recently, within functionally unrelated multi-domain proteins, such as pregnancy associated plasma proteins and stealth proteins. Prosite database defines LNR (PDOC50258) as a 35 amino acid module with three conserved Asp/Asn residues and six Cys residues engaged in a particular disulfide pattern favored by the presence of Ca2+. However, homology searches reveal naturally occurring LNRs with only four of the conserved Cys residues, as well as deviations in the proposed Ca2+ binding residues (Figure 1A). A. B. In this work we investigated the impact of free [Ca2+] and the total number of disulfides on the reduction of LNRs under various redox potentials through a comparative study of multiple LNRs. Our results indicate that while bound Ca2+ provides significant protection and prolonged chemical stability against reduction under even strong reducing conditions for the canonical LNRs, this Ca2+ based tunable stability is eliminated for LNRs missing the first pair of disulfide bonds, despite the presence of all the conserved Ca2+ binding residues. Taken together with our earlier findings we propose that LNRs are small protein modules that have evolved to provide varying amounts of redox sensitivity to the multi-domain protein they are incorporated in through a protein specific arrangement of multiple LNR modules with subtle, yet critical sequence variations. Results: Below the fully saturating Ca2+ concentration, the rate of hN1LNRA reduction is redox potential dependent Ca2+ protects the disulfide bonds against reduction and prolongs the chemical stability of folded hN1LNRA hN1LNRA:EEAC-ELPECQEDA-GNKVCSLQCNNHACGWDGGDC GS LNRA : EC—-AEGCPGSWIKDGYCDKACNNSACDWDGGDC hN4LNRA :GAKGCEGRS-GDGACDAGCSGPGGNWDGGDC Figure 1. (A) Sequence alignments of the first LNRs from three different proteins: Human Notch 1 (hN1LNRA), N-acetylglucosamine-1-phosphate transferase (GS LNRA), and Human Notch 4 (hN4LNRA). Cysteine residues are in orange, and the Ca2+ coordinating D/N residues in yellow. The characteristic disulfide bonding-pattern is indicated above the sequence. 97.5% LNRA-Ca2+ complex 99.8% LNRA-Ca2+ complex (B) NMR structure of hN1LNRA with disulfide bonds shown as orange sticks and Ca2+ coordinating residues represented as yellow sticks.1 Figure 4. Percent of reduced hN1LNRA, complexed with Ca2+ to varying degrees, after 60 min. of reaction with 250 μM (E=-338±6 mV) and 2.5 mM DTT (E=-361±1 mV). At Ca2+ concentrations below what is needed to form essentially 100% LNRA-Ca2+ complex, the amount of reduction at a given time was directly proportional to the reducing power of the environment. When hN1LNRA was fully saturated with Ca2+, however, it was fully protected against reduction at both redox potentials. N15 C4 C9 C22 Figure 3. Reduction of folded hN1LNRA as a function of time in varying [Ca2+] and 2.5 mM DTT. Percent values of reduced hN1LNRA in 0μM,25 μM,100 μM,1 mMand10 mMfree [Ca2+] were plotted as a function of time in 15 minute intervals up to 1 hr. Ca2+ C27 D30 D33 Conclusions: N-term S19 C34 C18 C-term • Ca2+ binding LNRs, like hN1LNRA and GS LNRA, are protected against reduction by free Ca2+ in the environment. The rate of reduction is inversely proportional to the free [Ca2+ ]. • The rate of reduction of an LNR is dependent on both the free [Ca2+ ] and the redox potential of the environment, unless the free [Ca2+ ] is fully saturating. At this point, there is complete protection against reduction for the range of tested redox potentials. • Ca2+ does not does not have an impact on the reduction of hN4 LNRA because hN4 LNRA does not bind Ca2+ . However, even in the absence of any Ca2+, hN4LNRA, with one fewer disulfide bond is still more susceptible to reduction at a given redox potential than its three disulfide-bonded homologs. Materials and Methods: Sample Preparation: Bacterially expressed hN1LNRA and synthesized GS and hN4LNRA were folded in a refolding buffer that allowed the formation of native disulfide bonds and were purified via reverse phase High Performance Liquid Chromatography (RP-HPLC). Folded proteins were then dialyzed against 20 mM HEPES, pH 8.0, 150 mM NaCl, containing predetermined concentrations of free Ca2+ as calculated by Visual Minteq.2 Reduction Reactions: Dialyzed proteins were aliquoted into reaction tubes to have a final concentration of 15 μM and were placed in an AtmosBag (Sigma-Aldrich) purged with N2. Dialysis buffers supplemented with 250 μM or 2.5 mM DTT were added under these anaerobic conditions to start the reduction reaction. At predetermined time points, reactions were quenched via acidification and assayed via analytical RP-HPLC, using a C18 column running a 0.1% / min. acetonitrile gradient. Data Analysis: The areas under each corresponding peak on the 280 nm chromatograms were integrated to quantitate the amounts of oxidized, reduced, and misfolded proteins (Figure 2). Percent oxidized, reduced, and misfolded proteins were calculated by dividing the integrated peak area of the corresponding peaks by the total area of all protein peaks from individual chromatograms (Table 1). Calculation of protein-Ca2+ complex percentage: The percent of protein complexed with Ca2+ at each free [Ca2+] was calculated using the formula3 ML = {Lo+Mo+KD) – ((Lo+Mo+KD)2 – 4MoLo)1/2} / 2, describing reversible binding between a receptor and a ligand (ML: % bound complex, Mo: initial [protein] = 15 μM, Lo: initial [Ca2+] of the experiments included in the tables, and Kd: dissociation constant, previously determined to be 25 μM). Calculation of Redox Potentials: To determine the amount of reduced and oxidized DTT present at each experimental time point, a set of experiments with known concentrations of oxidized and total DTT were performed in the absence of protein. Peaks on the chromatograms corresponding to the oxidized (only at 280 nm) and reduced (at 229 and 280 nms) were integrated to determine the corresponding absorbances. These values were substituted into Beer’s Law (A = εlc) to calculate the extinction coefficients of the oxidized and reduced forms of DTT. These extinction coefficients were used to determine the actual concentration values at the assayed time points of the reduction experiments. The redox potentials for each experiment and time point were calculated using the Nernst equation Eh (in mV) = Eo – (RT/nF) ln([DTTred]2/[DTToxid]); Eo = -323 mV at pH 7.0 with an adjustment of -5.9 mM / 0.1 increase in pH.4 Discussion: • Ca2+ is an integral part of the LNR structure possessing three disulfide bonds1. Fluorescence experiments have shown that binding of Ca2+ to the LNR alters the surface exposure of residues around the binding pocket. Hence the [Ca2+] dependent protection against reduction observed for these LNRs can be attributed to the increased chemical stability of the LNR-Ca2+ complex compared to its apo form due to a decrease in the solvent accessibility of the disulfide bonds upon Ca2+ binding. • In comparison to the cytoplasm, which has a resting redox potential of approximately -230 mV that can be significantly altered by cellular status (-240, -200, and -170 mV during cell proliferation, differentiation, and apoptosis, respectively), the ER lumen provides a relatively constant and more oxidative environment (-180 mV)4 critical for the proper folding of extracellular proteins. Unlike redox potential, though, the free [Ca2+ ] in the ER can change significantly based on cellular demands.6 • LNRs that are the focus of this study are found as repeated units within different multi-domain proteins targeted to the cell membrane. During folding in the ER, their disulfide bonds are formed and broken until the correct bonding pattern is achieved. The ability of free Ca2+ in the environment to selectively fine tune the chemical stability of the LNRs by altering their redox sensitivity offers a novel mechanism for the cells to regulate this process and preserve any correctly folded regions of the protein over misfolded regions, which can continue to shuffle their disulfide bonds in search of the most stable conformation. Table 2. Quantification of hN1LNRA reduction over time in 20 mM HEPES, pH 8.0, 150 mM NaCl, 2.5 mM DTT, and varying free [Ca2+]. The percent of reduced hN1LNRA and the redox potentials for each experiment were determined as described in the Materials and Methods section. Since the presence of metal ions impacts the stability of DTT5, the redox potentials calculated in the presence of different [Ca2+] were averaged for each time point, excluding the EDTA conditions, which are reported separately. Based on the calculated redox potentials, oxidized hN1LNRA experienced very similar reducing environments (redox potential = -343±6) in the presence of any amount of Ca2+ during the course of the experiment. Under these redox conditions, essentially full protection against reduction was achieved when there was enough free Ca2+ to ensure >97.5% LNRA-Ca2+ complex. Correctly folded hN1 LNRA peak (13.0 min.) Figure 2. Overlay of chromatograms recorded at 280 nm at 0, 15, 30, 60, 180min. for the series of hN1LNRA reduction experiments. The four sets of quantified peaks are annotated on the figure. Reduced hN1 LNRA peak (17.0 min.) Oxidized DTT peak (9.50 min.) Misfolded hN1LNRA Peaks Folded hN1LNRA and GS LNRA are more protected against reduction by Ca2+ than hN4LNRA Table 1. Integrated peak areas (IPA) of the corresponding peaks on the chromatogram, in addition to the calculated oxidized and reduced hN1LNRA percentages and redox potentials for 250 μM DTT in 0 μM Ca2+ (50 μM EDTA) at 0, 15, 30, 60, 180min. • References: • Vardar, D., North, C.L., Sanchez-Irizarry, C., Aster, J. C., Blacklow, S. C. (2003) Nuclear Magnetic Resonance Structure of a Prototype Lin12-Notch Repeate Module from Human Notch1. Biochemistry, 42, 7061-7067. • Visual MINTEQ: http://www2.lwr.kth.se/English/OurSoftware/vminteq/ • Jakubowski. “Chapter 5 – Binding. A: Reversible Binding 1 Equations and Curves.” (2010) http://employees.csbsju.edu/hjakubowski/classes/ch331/bind/olbindderveq.html. • Shafer, F.Q., & Buettner, G.R. (2001) Redox environment of the cell as viewed through the redox state of the glutathione disulfide/glutathione couple. Free Radical Biology & Medicine, 30(11), 1191-1212. • Burmeister Getz, E., Xiao, M., Chakrabarty, T., Cooke, R., & Slevin, P.R. (1999) A comparison between the sulfhydryl reductants tris(2-carboxyethyl)phosphine and dithiothreitol for use in protein biochemistry, Analytical Biochemistry, 273, 73-80. • Bygrave, F.L, & Benedetti, A. (1996) What is the concentration of calcium ions in the endoplasmic reticulum? Cell Calcium, 19 (6), 547-551. Table 3. Quantification of the three homologous LNRs over time in Ca2+-free and Ca2+-saturated environments. The percent of reduced hN1LNRA, GS LNRA, and hN4LNRA in 0 μM and 10 mM free Ca2+ and 250 μM DTT at 0, 15, 30, and 60 minutes, as well as the redox potentials calculated as described in the Materials and Methods section. hN4LNRA, with one fewer disulfide bond, has a faster rate of reduction in comparison to hN1LNRA and GS LNRA. The varying concentrations of free Ca2+, in fact, had no effect in the reduction of hN4LNRA, while the saturation of Ca2+ significantly protected and prolonged the chemical stability of hN1LNRA and GS LNRA against reduction. • Funding: • Camille and Henry Dreyfus Faculty Start-up Award (DVU) • Wellesley College Sophomore Early Research Program (JJ, AS) • Protein Science Young Investigator Travel Grant (JJ) • Wellesley College Science Center Travel Award (JJ, AS)