Download

1 / 17
780 likes | 2.48k Views
Cellular protein degradation. 17-1. Proteolytic pathways in eukaryotes lysosomal degradation of proteins ubiquitin-proteasome dependent protein degradation post-proteasome degradation: Tricorn, TPII membrane protein degradation. Main proteolytic pathways in eukaryotes. 17-2.

E N D
Cellular protein degradation 17-1 • Proteolytic pathways in eukaryotes • lysosomal degradation of proteins • ubiquitin-proteasome dependent protein degradation • post-proteasome degradation: Tricorn, TPII • membrane protein degradation
Main proteolytic pathways in eukaryotes 17-2 • endosome-lysosome pathway degrades extracellular and cell-surface proteins • ubiquitin-proteasome pathway degrades proteins from the cytoplasm, nucleus and ER • mitochondria (and chloroplasts) have their own proteolytic system of bacterial origin Mitochondrial proteolytic system cytoplasmic proteins endosome- lysosome system Ubiquitin- proteasome system Lysosome/ endosome Mitochondria cooperation? Autophagosome ER proteins nuclear proteins Nucleus
Timeline of regulated intracellular proteolysis 17-3 Schoenheimeruses 15N to show continuous protein turnover Hershko et al. identify enzymes of the ubiquitin-protein ligase system Kischner et al. discover that cyclin is degraded by the ubiquitin pathway 26S proteasome partly purified by Rechsteiner Folin states “endogenous proteins are stable” Aaron Ciechanover Avram Hershko Irwin Rose Nobel Prize 2004 1905 1912 1942 1978 1983 1984 1986 1988 1991 1997 Lewis discovers ‘bodies’ in patients with Parkinson’s disease Hershko and Ciechanover discover the process of ubiquitylation Varshavsky, Ciechanover and Finley discover ubiquitylation is essential for viability and cell-cycle progression Lowe, Landon and Mayer discover that Lewy bodies are full of ubiquitylated proteins Several groups discover the combinatorial control and specificity of SCF ubiquitin protein ligases ubiquitin structure 1987 proteasome structure 1995 adapted from R. John Mayer, Nature reviews1, 145-148.
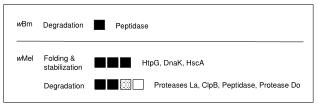
Lysosomal degradation of proteins 17-4 • lysosomes are cellular vesicles containing proteolytic enzymes (e.g., papain-like cysteine protease, serine proteases, aspartic proteinases, etc., which are typically monomeric • pH maintained at ~5.5 by proton-pumping ATPase • account for 1-15% of cell volume (most abundant in liver and kidney) • Most lysosomal enzymes are transported to lysosomes through recognition by receptors for mannose-6-phosphate. Lysosomal enzymes are synthesized like proteins destined to be secreted or for residence on the plasma membrane but are recognized by a phosphotransferase enzyme shortly after leaving the ER. This enzyme transfers N-acetylglucosamine-1-phosphate to one of more mannose residues. A glucosaminidase next removes the glucosamine to generate the M6P. • a mutation in the transferase leads to disease (I-cell disease); other so-called lysosomal storage diseases are the Tay-Sachs syndrome (ganglioside accumulates due to beta-Hexosaminidase deficiency), Pompe disease (accumulation of glycogen due to lack of -Glucosidase), etc. (6 others!)
Lysosomal degradation of proteins 17-5 • macroautophagy is the equivalent of forming intracellular endosomes (phagosomes) that fuse to the lysosome and result in the breakdown of its contents • Hsc73 (constitutively-expressed Hsp70 chaperone) is involved in one pathway of lysosome-mediated degradation Cuervo and Dice (1998) J. Mol. Med.76, 6-12.
The ubiquitin degradation pathway 17-6 • E1 -ubiquitin activating enzyme • E2 - ubiquitin conjugating enzyme • E3 - ubiquitin ligase • ‘~’ denotes high-energy thioester bond • DUB, deubiquinating enzyme
Ubiquitin-mediated degradation O O O O ub ub ub ub AMP S~E1 OH S~E2 17-7 • E1 -ubiquitin activating enzyme • uses ATP to activate the carboxyl group of ubiquitin’s C-terminal residue (Gly76). The outcome of this reaction is the formation of a thioester between Gly76 of ubiquitin, and a cysteine residue of E1 • E2 - ubiquitin conjugating enzyme • accepts the ubiquitin from the E1 through a thioester linkage with a cysteine • E3 - ubiquitin ligase • transfers the ubiquitin molecule to the epsilon NH2 group of lysine on the substrate • ubiquitin molecules are then added in succession to the Lysine 48 residue to form a multiubiquitin chain • the DUB enzyme ‘recycles’ ubiquitin • the 26S proteasome degrades the substrate to peptides E1-SH ATP E1-SH O ub N-H prot E2 E3 prot
17-7b E3 ligase
E3 ubiquitin ligases 17-8 • there are two basic types of E3 ubiquitin ligases: • those possessing Ring fingers (e.g., VHL, SCF, APC, MDM2, c-CBL, etc.) • those possessing HECT domains (E6AP-related proteins) VHL/SOCS-box SCF (Skp1/Cul/F-box) • Shown here are VHL and SCF ubiquitin ligases. They both associate with Rbx-1, an evolutionarily conserved protein containing a so-called ‘ring finger’ (not shown in figure) • ring fingers are also present in other ligases such as the APC and MDM2, which is involved in ubiquitinating p53 • CDC34 is modified with the ubiquitin-like protein rub-1; ElonginB also has homology with ubiquitin SH2, WD40, Ank, LRR are all protein-protein interaction domains
E3 ubiquitin ligase: VHL-Elongin B/C 17-9 • the domain serves to target proteins for degradation; HIF (hypoxia inducible factor) is one of the targets • VHL mutations cause tumours (VHL surface mapped with common mutations): crystal structure of a core ubiquitin ligase Stebbins et al. (1999) Science 284, 455-461.
c-CBL E3/E2/kinase structure 17-10 E3 E3 • c-Cbl proto-oncogeneis a RING family E3 that recognizes activated receptor tyrosine kinases (e.g., ZAP-70), promotes theirubiquitination by a ubiquitin-conjugating enzyme (E2) and terminates signaling • crystal structure of c-Cblbound to a cognate E2 and a kinase peptide shows how theRING domain recruits the E2. A comparison with a HECT family E3-E2 complexindicates that a common E2 motif is recognized by the two E3families E2 substrate E3 E3 Zheng et al. (2000) Cell 102, 533-539.
SCF-dependentubiquitinationin yeast 17-11 • F-box proteins mediate substrate selectivity in degrading various yeast proteins • many (all?) of the substrates need to be phosphorylated to be recognized by the F-box protein • WD40 and leucine-rich repeats (LRRs) present in F-box proteins mediate protein-protein interactions
Anaphase promoting complex (APC) 17-12 • Theanaphase-promoting complex (also termed ‘cyclosome’) is a ubiquitin-protein ligase thatcontrols important transitions in mitosis by ubiquitinatingregulatory proteins • consists of many different proteins, including some related to SCF (e.g., ring protein) • To initiatesister chromatid separation, the APC has to ubiquitinatethe anaphase inhibitor securin, whereas exit from mitosisrequires the ubiquitination of B-type cyclins em reconstruction unprocessed em images Gieffers et al. (2001) Mol. Cell7, 907-913.
Tricorn protease of prokaryotes 17-13 • tricorn protease is a huge hexameric protease complex that assembles into even larger cage-like structure containing 20 hexamers (14.6 MDa) • cage required for efficient degradation? example of self-compartmentalization void volume! huge cryo-em reconstruction of tricorn capsids (A) tricorn protease exists as 2 different species; one of ~730 kDa and one much larger which elutes in the void volume of the sizing column (B) electron microscopy (em) of the 730 kDa species Walz et al. (1997) Mol. Cell1, 59-65.
Tricorn protein degradation pathway 17-14 • tricorn protease in prokaryotes may be part of a degradation pathway that involves proteasome (in archaea) or other ATP-dependent proteases in archaea/bacteria • proteasomes/other oligomeric proteases digest proteins to small peptides • tricorn protease then cleaves these to 2-4 mers, which are then degraded down to the level of free amino acids by aminopeptidases a modular system for protein degradation • probably one of many pathways of protein degradation in prokaryotes Yao and Cohen (1999) Curr. Biol.9, 551-553.
Tricorn-like protease in eukaryotes? 17-15 • tripeptidyl peptidase II (TPPII) is acytosolic subtilisin-like peptidase that may be functionally related to Tricorn protease • discovery • cells adapted to near-lethal concentrations of vinyl sulphone(VS)-proteasome inhibitors still have the ability to degrade ubiquitinated proteins, control the cell cycle, and present MHC class I peptides • cells had alanyl-alanyl-phenylalanyl-7-amino- 4-methylcoumarin(AAF-AMC)-hydrolyzing activity in size exclusion fractions larger than proteasome • forms on average tripeptides • general proteolytic activity em pictures of TPPII; dumbbell- or ovoid-shaped Legend to gels: Galactosidase (116 kD) (lane 1), purified TPPII (lane 2), fast- and slow-running electrophoretic isoforms of 26S proteasomes (lanes 3 and 4, respectively), and purified 20S proteasomes (lane 5) Geier et al. (1999) Science283, 978-981.
Membrane protein degradation 17-16 • AAA proteases mediate the degradation of membrane proteins in bacteria, mitochondria and chloroplasts (i.e., compartments of eubacterial origin) • bacterial Lon, FtsH combine proteolytic and chaperone activities in one system, acting as quality-control machineries -model substrate polypeptides containing hydrophilic domains at either side of the membranecan be completely degraded by either of two AAA proteases found in mitochondria, if solvent-exposed domainsare in an unfolded state - a short protein tail protruding from the membranesurface is sufficient to allow the proteolytic attack of an AAA proteasethat facilitates domain unfolding at the opposite side wt-DHFR stabilises protein degradation degradation p=precursor; m=mature; 25ºC=no unfolding;37ºC=unfolding of domain(s) Leonhard et al. (2000) Mol. Cell 5, 629-638.