Download

1 / 66
660 likes | 856 Views
Protein-Protein Interactions and Inhibition of the ADP-Ribosyl Transferase Reaction of Pseudomonas aeruginosa Exotoxin A. Susan P. Yates Ph.D. Thesis Defence Supervisor: Dr. A. Rod Merrill. Outline. Background Research Objectives Inhibition of the catalytic domain of exotoxin A

E N D
Protein-Protein Interactions and Inhibition of the ADP-Ribosyl Transferase Reactionof Pseudomonas aeruginosa Exotoxin A Susan P. Yates Ph.D. Thesis Defence Supervisor: Dr. A. Rod Merrill
Outline • Background • Research Objectives • Inhibition of the catalytic domain of exotoxin A • Interactions between the toxin and its protein substrate • Final Thoughts
Pseudomonas aeruginosa • Gram-negative rod-shaped bacterium • Opportunistic pathogen • Exploits some break in the host defenses to initiate an infection • Cystic fibrosis, severe burns, AIDS, cancer, etc. • Highly adaptable to new environments • Resistant to many antibiotics • Possesses a vast array of virulence factors • Very complex pathogenesis
Virulence Factors Pilus Flagellum Pseudomonas aeruginosa Alginate/Biofilm LPS • Extracellular products • Rhamnolipid • Phospholipase C • Proteases • Siderophores • . Exotoxin A
Exotoxin A – The Virulence Factor • Exotoxin A (ETA) is the most potent virulence factor of Pseudomonas aeruginosa • LD50 of 0.2 mg when injected intraperitoneally into a 18-gram mouse • Biological effects • Extensive tissue damage • Promotes bacteria invasion • Interferes with function of the cellular immune system • May lead to systemic disease
Exotoxin A – The Enzyme • Member of mono-ADP-ribosyl transferase family • Other members include: • Diphtheria toxin, pertussis toxin, cholera toxin, C3 exoenzyme, iota toxin • 66 kDa single polypeptide • Three functional domains • Secreted as a proenzyme • Activated within the eukaryotic cell through a proteolytic event III Catalytic Ib II Translocation Ia Receptor binding (Wedekind et al., (2001) J. Mol. Biol.314, 823)
Eukaryotic Elongation Factor 2 (eEF2) • Protein substrate for ETA • 90 –110 kDa protein • GTPase superfamily • Important factor in the elongation step of protein synthesis • Covalent modification by ETA produces ADP-ribosyl eEF2 (ADPR-eEF2) • Prevents its participation in protein translation • Cell death G′ G II III V IV Diphthamide (Jørgensen et al., (2003) Nat. Struc. Biol.10, 379)
Catalytic Domain of ETA (PE24H) (Li et al., (1996) PNAS93,6902)
Research Objectives – The Big Picture • General statement • Improve the understanding of the interactions between the catalytic domain of ETA and both its substrates, eEF2 and NAD+ • Long term research goals • Understand the detailed reaction mechanism for ETA • Knowledge-based approach to preventing the action of this toxin • Develop new strategies that target ETA to fight Pseudomonas aeruginosa infections
Research Objectives – My Specific Projects Part A: Interactions of the toxin with NAD+ • Study of water-soluble inhibitors • Development of a NAD+-glycohydrolase assay Part B: Toxin-eEF2 interactions • Physiological requirements for binding • Fluorescence-based approach to elucidate sites of contact • Fluorescence resonance energy transfer (FRET) distance study
PART A: Interactions of the Toxin with NAD+ Project #1 STUDY OF WATER-SOLUBLE INHIBITORS Yates, S.P., Taylor, P.L., Jørgensen, R., Ferraris, D., Zhang, J., Andersen, G.R., and Merrill, A.R. Biochem. J. (2005) 385:667-675.
Inhibition of PE24H • Previous work from our research group • Characterization of a series of small, non-polar competitive inhibitors • Most potent inhibitor was NAP (1,8-napthalamide) • Model of NAP bound to catalytic domain of ETA • Lack of water-solubility limited the usefulness as potential therapeutic drugs Armstrong et al., (2002) J. Enzyme Inhib. Med. Chem. 17, 235
Aims of Study • Characterize a series of water-soluble compounds for their inhibition against PE24H • Co-crystal structure of the inhibitor PJ34 with PE24H
The Inhibitors • Mimic nicotinamide • IC50 values ranged from 170 nM to 82.4 mM • GP-D, PJ34, GP-M most potent • Hallmark of a good inhibitor was a planar hetero-ring
PJ34 – Further Characterized • Water-soluble phenanthridinone derivative • IC50 = 280 nM • Commercially available • Well-characterized compound • Studied in extensively in several PARP related systems
Biochemical Characterization of PJ34 • Binding affinity • KD is 820 54 nM • 70x tighter binding to PE24H compared to NAD+ • Competitive inhibitor • As [PJ34] increases, the KM increases but the Vmax remains unchanged • Ki = 140 nM determined using both Dixon and Lineweaver-Burk methods
Crystallization of PE24H-PJ34 • Data • 2.1 Å resolution • Refinement • R-factor = 21.3 % • Rfree-factor = 23.5 %
Hydrophobic Pocket and Active Site Yates et al., (2005) Biochem. J. 385, 667
Interactions in the Active Site 3.1 Å 2.5 Å 2.7 Å 2.5 Å Yates et al., (2005) Biochem. J. 385, 667
Similar Enzymes • Catalytic domain of ETA is functionally and structurally similar to both mono-ADPRTs and PARPs • Diphtheria toxin (DT) • Mono-ADPRT and also catalyzes the ADP-ribosylation of eEF2 • PARPs (Poly-(ADP-ribosyl) polymerases) • Catalyzes the covalent attachment of ADP-ribose units to nuclear DNA-binding proteins Taken from: Putt & Hergenrother (2004) Anal. Biochem.326, 78
Comparison to Other Active Sites DT DT structure: Bell & Eisenberg, (1996) Biochemistry35, 1137 PARP PARP structure: Ruf et al., (1998) Biochemistry37, 3893
Findings for Project #1 • Hetero-ring planarity important for inhibition • PJ34 is a competitive inhibitor • First report of a structure of a mono-ADPRT-inhibitor complex • Confirmed the hydrogen bonding of the lactam moiety to Gly-441 • Planar compounds sandwich better into the nicotinamide-binding pocket than more flexible compounds • Similarities and differences between bacterial toxins and PARP • Exploit the differences to target one enzyme over the other
PART A: Interactions of the Toxin with NAD+ Project #2 DEVELOPMENT OF A NAD+-GLYCOHYDROLASE ASSAY Yates, S.P., and Merrill, A.R. Anal. Biochem. (2005) in press.
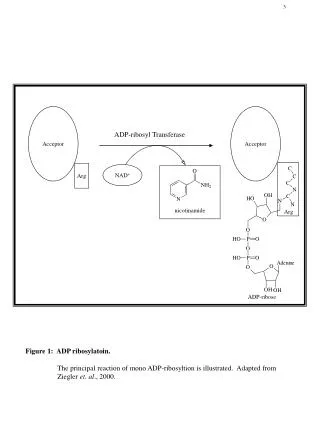
NAD+-Glycohydrolase Activity H N 2 N O N O O N NH2 + N N H C P P CH2 2 O O O O O - - O O H H O OH H O OH NAD+ O N H - 2 STEP 1 H N 2 N N nicotinamide N O O N N H C P P CH2 2 + O O O O O - - H O O - + O H + H H O OH H O OH oxacarbenium ion O H 2 H N + STEP 2 - H 2 N N O O N CH2 H N H C P P O 2 O O O O - - O O OH H O OH H O OH ADP-ribose
F-NAD+ • Initial inhibitor study showed that IC50 value is 82.4 7.4 M • Binding affinity to toxin similar to NAD+ • NAD+ KD = 53 2 M • F-NAD+ KD = 33 1 M
Aims of Study • Is F-NAD+ a competing substrate or a competitive inhibitor? • Is the C-N bond broken? • Develop an HPLC-based NAD+-glycohydrolase assay • Why? • Fluorometric assay uses e-NAD+ • Contains a etheno bridge which gives rise to its fluorescence • F-NAD+ lacks this structural feature
Reaction and Sample Preparation Samples (25 mL) taken at t = 0 to 4 hrs Reaction Setup Toxin + NAD+ (250 mL) Sampling Inhibit Reaction To HPLC PE24H bound to resin Load to Spin Column Toxin Removed Add 75 mL Mobile Phase (with internal standard) Flow-Through ready for HPLC – contains no protein Chelating Sepharose Spin Column
HPLC Instrumentation Setup Inject sample via sample loop 150 mm C18 column – reverse phase Precolumn 4.6 mm Detector at 259 nm Mobile phase: 20 mM NaHPO4, pH 5.5: acetonitrile (100:5 v/v %)
HPLC and Analysis – Rate Determination Rate = 55 3 mM nicotinamide produced per hour + Nicotinamide Standard Curve Chromatogram Time Course Plot
Rate of Hydrolysis of F-NAD+ • Visual inspection of chromatograms shows the peak area for ADPR increasing • Mathematically deconvoluted ADPR peak from NAD+ or F-NAD+ peak • Hydrolysis of F-NAD+ is 0.2% rate of NAD+
Findings for Project #2 • HPLC-based NAD+-glycohydrolase assay developed • Addition of spin column step allows quick removal of protein • F-NAD+ binds to the enzyme but not readily hydrolyzed • What does fluorine substitution at 2'-OH position do? • Disrupts hydrogen bond between Glu-553 and 2'-OH position • This hydrogen bond important for bond breakage • Cause nicotinamide leaving group to depart slower • Fluorine substituent may destabilize cationic intermediate
PART B: Toxin-eEF2 Interactions Project #3 PHYSIOLOGICAL REQUIREMENTS FOR BINDING Loop Yates, S.P., and Merrill, A.R. J. Biol. Chem. (2001) 276:35029-35036. pH and Guanyl nucleotide Armstrong, S., Yates, S.P., and Merrill, A.R. J. Biol. Chem. (2002) 277:46669-46675. ADPR-eEF2 Jørgensen, R., Yates, S.P., Teal, D.J., Nilsson, J., Prentice, G.A., Merrill, A.R., and Andersen, G.R. J. Biol. Chem. (2004) 279:45919-45925.
Aims of Study • Investigate the conditions required for toxin-eEF2 interaction • Effect of pH • Effect of bound guanyl nucleotides on eEF2 • Effect of ADP-ribosylation of eEF2 • Functional role of a surface-exposed loop near the active site
FRET-based eEF2 Binding Assay • Fluorescence Resonance Energy Transfer (FRET) • Transfer of excitation energy from a donor fluorophore to a an acceptor fluorophore through non-radiative dipole-dipole interactions • Criteria • Donor and acceptor in close proximity • Acceptor absorption overlaps with fluorescence emission of donor • Dipole-dipole interactions are parallel • Donor fluorophore • PE24H labelled with IAEDANS (PE24H-AEDANS) • Acceptor fluorophore • eEF2 labelled with fluorescein (eEF2-AF)
Effect of pH on eEF2 Binding to Toxin • Optimum eEF2 binding at pH 7.8 • Two distinct pKa values • Acidic pKa = 6.3 • His residue • Alkaline pKa = 9.3 • Tyr residue • pH profiles for eEF2 binding and catalysis very similar • eEF2 binding may be responsible for pH dependence observed in catalysis
Effect of Guanyl Nucleotides • eEF2 is a member of the GTPase superfamily • Does the toxin require a specific eEF2 conformation for binding? • eEF2 with non-hydrolyzable GTP/GDP analogues bound • Toxin does not prefer a specific state of eEF2 for either binding or catalytic function
Interaction of ADPR-eEF2 with Toxin • ADPR-eEF2 maintained the ability to bind toxin • Active site of toxin can accommodate the bulky ADP-ribose group • Structures of both eEF2 and ADPR-eEF2 recently solved • No major conformational changes induced after ADP-ribosylation
Characterization of a Loop in ETA • History of Loop C • Residues 483-490 • Functional removal • Decreases activity significantly (1.8 x 10+4-fold) • Retains ability to bind NAD+ near wild-type levels • Alanine-scanning mutagenesis • Some mutant proteins exhibited reduced activity • KD and KM for NAD+ similar to wild-type • What is the role of this Loop? • Catalytic or eEF2 substrate binding? Loop C (Li et al., (1996) PNAS93,6902)
Determination of KM and KD for eEF2 • pG-Loop C mutant protein • Each residue within Loop C replaced with glycine • Functional removal of loop • Retained ability to associate with eEF2 at normal levels • Alanine-scanning mutants • KM for eEF2 unaffected • Enzyme rate (kcat) is affected • Loop is a catalytic element • May modulate the transferase activity of the toxin
Findings for Project #3 • Toxin-eEF2 association is pH-dependent • Correlates to that observed for catalytic function • GTP or GDP bound to eEF2 did not affect it as a protein substrate • Structurally the diphthamide and guanyl nucleotide binding site are quite distant • No direct coupling of sites • Toxin maintains the ability to associate with eEF2 after its ADP-ribosylation • Loop C is important for catalysis • May stabilize the transition state structure during the catalytic reaction
PART B: Toxin-eEF2 Interactions Project #5 FLUORESCENCE-BASED APPROACH TO ELUCIDATE SITES OF CONTACT Yates, S.P., and Merrill, A.R. Biochem. J. (2004) 379:563-572.
Aim of Study • Identifycontact sites between eEF2 and PE24H • This protein-protein interaction is poorly characterized • Two extreme models are possible • Minimal Contact Model Maximum Contact Model PE24H eEF2 eEF2 PE24H
Experimental Approach • Single cysteine residuesintroducedinto PE24H at 21 defined surface sites andlabelledwith the fluorophore, IAEDANS O IAEDANS I NHCH CH NH C CH 2 2 2 .. CH PROTEIN HS 2 SO H 3 O NHCH CH NH S CH PROTEIN C CH 2 2 2 2 Protein adduct (Li et al., (1996) PNAS93,6902) + HI SO H 3
Experimental Approach • Fluorescence studies performed in the presence and absence of eEF2 • Fluorescence wavelength emission maxima (lem,max) • Fluorescence lifetime • Acrylamide quenching
Acrylamide Quenching F0/F 1 [Q] • Measure the ability of acrylamide to quench the fluorescence of IAEDANS probe attached to PE24H • Acrylamide is a water-soluble, non-ionic quencher • The more accessible the probe is to acrylamide, the more quenching is observed • Determine the bimolecular quenching constant (kq) in the presence and absence of eEF2 using the Stern-Volmer equation • kq is the rate of collisions with the quencher that result in deactivation of excited state of the fluorophore
Crude Model of PE24H-eEF2 Complex Domain IV Domain IV of eEF2 of eEF2 diphthamide diphthamide 519 3 7 486 459 4 554 442 449 6 5 410 8 2 507 408 1 9 PE24H PE24H • Potential eEF2 contact sites on PE24H • Minimal contact between proteins • Diphthamide residue on eEF2 positioned near scissile glycosidic bond of NAD+ in active site (Li et al., (1996) PNAS93,6902; Jørgensen et al., (2003) Nat. Struc. Biol.10,379)