Download

1 / 15
150 likes | 199 Views
Explore the intricate process of cellular respiration in non-photosynthetic cells, detailing how high-energy compounds are utilized to generate ATP. Learn about the role of mitochondria, electron transport chain, and ATP synthesis.

E N D
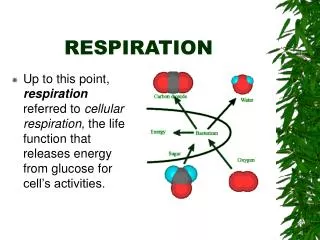
Respiration Also non-photosynthetic cells need energy. They ‘burn’ high energy compounds (‘food’) to generate the energy they need via a process called ‘respiration’. In eukaryotic cells respiration occurs in the mitochondrion. Interestingly, mitochondria (like chloroplasts) carry their own DNA, suggesting a complex evolutionary origin. In essence the process of respiration proceeds as follows: food is chemically modified in reduced compounds (NADPH, sugars) which serve as an electron donor. The respiratory chain consists of a set of membrane bound proteins, via which the electron is transported to reduce oxygen (that we breathe) to water. During this electron transport reaction protons are transported across the mitochondrial membrane to build up a proton gradient. The proton gradient is used by the enzyme ATPsynthase to produce ATP, the universal energy compound of the living cell.
Creating ATP Fig 4.36 The general mechanism of oxidative phosphorylation. As a high energy electron is passed along the electron transport chain, some of the energy that is released, is used to drive the three respiratory enzyme complexes that pump H+ out of the matrix. The resulting electro-chemical proton gradient across the inner membrane drives H+ back through the ATP-synthase, a membrane protein complex that uses the energy of the H+ flow to synthesize ATP from ADP and Pi in the matrix.
ATP to ADP The dephosphorylation reaction of ATP to ADP and inorganic phosphate is often coupled with nonspontaneous reactions to drive them forward Nature uses ATP as a free energy currency, energy liberated ~ 20kBT thermal energy <ATP < covalent bond (150 kBT) Also the production of ATP from ADP is coupled to a spontaneous reaction:
Production of ATP PhosphorylationADP3- + HPO42- + H+ -->ATP4- + H2O ΔGo= +30.5 kJ(nonspontaneous) oxidationNADH --> NAD+ + H+ + 2e-ΔGo= -158.2 kJ(spontaneous) reduction1/2 O2 + 2H+ + 2e- --> H2OΔGo= -61.9 kJ(spontaneous) In cells these reactions are coupled via a proton-pumping system
Cytochrome c oxidase Cytochrome c docking site O2 is trappedbetween heme-Fand Cu until it isfully reduced toH2O O2- is dangerous e- Propagation by diffusion is the fastest mode of mass transfer over very short distances, Proton through gramicidin channel ~15 ns Block channel from becoming a proton leak!
The photosynthetic pigments An extensive delocalized π electron system extends over most of the molecule, with the exception of ring D, in which the C-17-C-18 double bond is reduced to a single bond. In BChl also ring B is reduced.
Spectroscopic Properties of Chlorophylls The absorption bands arise from π->π* transitions, involving the electrons in the conjugated π system of the chlorin macrocycle.
Absorption (left) and fluorescence (right) spectra of chlorophyll and bacteriochlorophyll a in diethyl ether. Note the 100 nm redshift of the spectrum and the enhanced splitting between the Qy and Qx transitions. Both are a consequence of the further loss of symmetry in bacteriochlorophyll a as compared to chlorophyll a. The extinction coefficient of bacteriochlorophyll a around 770 nm is about 100 mM-1cm-1
MO level diagram The diagram is an oversimplification of what is a complex relationship between electronic states and orbital energies. It suggests that electronic transitions reflect a simple promotion of an electron from a HOMO to a LUMO. In reality, several different electronic configurations, including contributions from much higher-energy MO’s, can contribute to the electronic transition. This phenomenon is known as configuration interaction. The result is that there is not a simple one-to-one correspondence between orbital occupations and electronic transitions.
Electronic transitions The electronic transitions are associated to transition dipole moments with different strengths and orientations. The longest-wavelength transition is invariably polarized along the y-axis of the molecule and is therefore known as the Qy transition. This means that the absorption will be strongest if the electric field vector of linearly polarized light is parallel to the y molecular axis of the pigment. The exciting light couples to the π-electrons of the molecule and transiently arranges them somewhat during the transition. The Qy transition causes a shift in electron density that is directed along the y molecular axis of the molecule Electronic density changes associated with Qx and Qy transitions in bacteriochlorophyll a. Note that the signs are chosen arbitrarily, the charge density changes oscillate with the incident light field.
Carotenoids • Collection of light • Photoprotection. • Regulation of energy transfer in the antennas.
Absorption of light by a C=C double bond excites a π electron into an antibonding π* orbital (fig.6.8). The chromophore activity is therefore due to a π -> π * transition. Its energy is about 7 eV for a single conjugated double bond (like in ethene), which corresponds to an absorption at 180 nm (in the ultraviolet). When the double bond is part of a conjugated chain (like in a carotenoid), the energies of the molecular orbitals lie closer together and the p->p* transition moves to longer wavelength; it may even lie in the visible region if the conjugated system is long enough. An important example of a p->p* transition is provided by the pigment retinal that plays the essential role in the photochemical mechanism of vision (see 6.1.7). When a carbonyl group (>C=O) is present (like in the peptide bond of proteins) an absorption at about 290 nm is normally observed, although its precise location depends on the nature of the rest of the molecule (a-helix vs b-sheet vs random coil).