Download
1 / 1
10 likes | 124 Views
Activin/Inhibin in Fundulus heteroclitus Ovary: Cloning of the Subunits and Effects on Oocyte Maturation . Program No. 238.17. School of Natural and Health Sciences, Barry University, Miami Shores, Florida 33161. Germinal Vesicle Prophase I. GVBD Metaphase II.

E N D
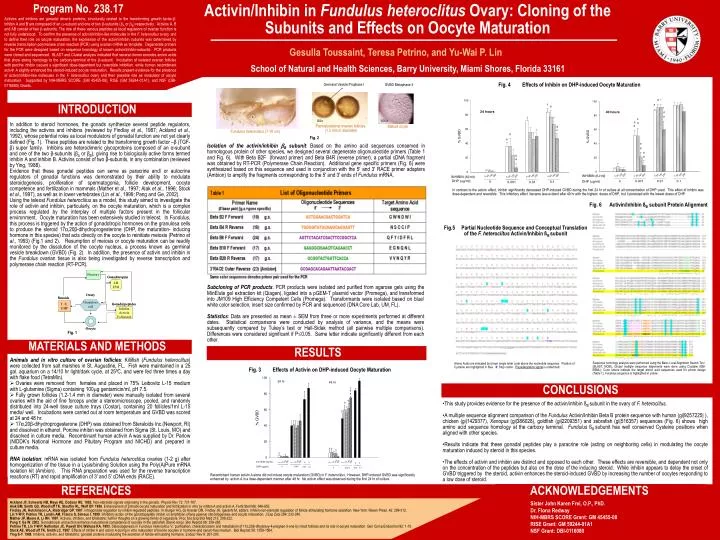
Activin/Inhibin in Fundulus heteroclitus Ovary: Cloning of the Subunits and Effects on Oocyte Maturation Program No. 238.17 School of Natural and Health Sciences, Barry University, Miami Shores, Florida 33161 Germinal Vesicle Prophase I GVBD Metaphase II Fig. 4 Effects of Inhibin on DHP-induced Oocyte Maturation 60x 100x Prematurational ovarian follicles (1.3 mm in diameter) Mature ocyte Fundulus heteroclitus (7-10 cm) Fig. 6 Activin/Inhibin ßB subunit Protein Alignment Fig.5 Partial Nucleotide Sequence and Conceptual Translation of the F. heteroclitus Activin/Inhibin ßB subunit MATERIALS AND METHODS Amino Acids are indicated by brown single letter code above the nucleotide sequence. Position of Cysteine are highlighted in blue. X: Stop codon. Polyadenylation signal is underlined. Sequence homology analysis were performed using the Basic Local Alignment Search Tool (BLAST, NCBI); Global multiple sequence alignments were done using Clustalw (EBI-EBML); Color letters indicate the target amino acid sequences used for primer design (Table 1), Fundulus sequence is highlighted in yellow c b a a b c a Activins and inhibins are gonadal dimeric proteins, structurally related to the transforming growth factor-. Inhibin A and B are composed of an -subunit and one of two -subunits (A or B) respectively. Activins A, B and AB consist of two -subunits. The role of these various peptides as local regulators of ovarian function is not fully understood. To confirm the presence of activin/inhibin-like molecules in the F. heteroclitus ovary, and to define their role on oocyte maturation, the expression of the activin/inhibin subunits was determined by reverse transcription-polymerase chain reaction (PCR) using ovarian mRNA as template. Degenerate primers for the PCR were designed based on sequence homology of known activin/inhibin-subunits. PCR products were cloned and sequenced. BLAST and Clustal analysis indicated that several clones encodes amino acids that show strong homology to the carboxy-terminal of the -subunit. Incubation of isolated ovarian follicles with porcine inhibin caused a significant dose-dependent but reversible inhibition, while human recombinant activin A slightly enhanced the steroid-induced oocyte maturation. Results present evidence for the presence of activin/inhibin-like molecules in the F. heteroclitus ovary and their possible role as modulator of oocyte maturation. Supported by NIH-MBRS SCORE (GM 45455-08), RISE (GM 59244-01A1), and NSF (DBI-0116080) Grants. Gesulla Toussaint, Teresa Petrino, and Yu-Wai P. Lin INTRODUCTION In addition to steroid hormones, the gonads synthesize several peptide regulators, including the activins and inhibins (reviewed by Findlay et al., 1987; Ackland et al.,1992), whose potential roles as local modulators of gonadal function are not yet clearly defined (Fig. 1). These peptides are related to the transforming growth factor –β (TGF- β) super family. Inhibins are heterodimeric glycoproteins composed of an α-subunit and one of the two β-subunits (βA or βB), giving rise to biologically active forms termed inhibin A and inhibin B. Activins consist of two β-subunits, in any combination (reviewed by Ying, 1988). Evidence that these gonadal peptides can serve as paracrine and/ or autocrine regulators of gonadal functions was demonstrated by their ability to modulate steroidogenesis, proliferation of spermatogonia, follicle development, oocyte competence and fertilization in mammals (Mather et al., 1997;Alak et al., 1996;Stock et al., 1997), as well as in lower vertebrates (Lin et al., 1999;Pang and Ge, 2002). Using the teleost Fundulus heteroclitus as a model, this study aimed to investigate the role of activin and inhibin, particularly, on the oocyte maturation, which is a complex process regulated by the interplay of multiple factors present in the follicular environment. Oocyte maturation has been extensively studied in teleost. In Fundulus, this process is triggered by the action of gonadotropic hormones on the granulosa cells to produce the steroid 17α,20β-dihydroprogesterone (DHP, the maturation- inducing hormone in this species) that acts directly on the oocyte to reinitiate meiosis (Petrino et al., 1993) (Fig.1 and 2). Resumption of meiosis or oocyte maturation can be readily monitored by the dissolution of the oocyte nucleus, a process known as germinal vesicle breakdown (GVBD) (Fig. 2). In addition, the presence of activin and inhibin in the Fundulus ovarian tissue is also being investigated by reverse transcription and polymerase chain reaction (RT-PCR). Fig. 2 Isolation of the activin/inhibin B subunit: Based on the amino acid sequences conserved in homologous protein of other species, we designed several degenerate oligonucleotide primers (Table 1 and Fig. 6). With Beta B2F (forward primer)and Beta B4R (reverse primer), a partial cDNA fragment was obtained by RT-PCR (Polymerase Chain Reaction). Additional gene specific primers (Fig. 6) were synthesized based on this sequence and used in conjunction with the 5' and 3' RACE primer adapters (Ambion) to amplify the fragments corresponding to the 5' and 3' ends of Fundulus mRNA. In contrast to the activin effect, inhibin significantly decreased DHP-induced GVBD during the first 24 hr of culture at all concentration of DHP used. This effect of inhibin was dose-dependent and reversible. This inhibitory effect became less evident after 40 hr with the highest doses of DHP, but it persisted with the lowest doses of DHP. Subcloning of PCR products: PCR products were isolated and purified from agarose gels using the MinElute gel extraction kit (Qiagen), ligated into a pGEM-T plasmid vector (Promega), and transformed into JM109 High Efficiency Competent Cells (Promega). Transformants were isolated based on blue/ white color selection, insert size confirmed by PCR and sequenced (DNA Core Lab, UM, FL). Statistics: Data are presented as mean ± SEM from three or more experiments performed at different dates. Statistical comparisons were conducted by analysis of variance, and the means were subsequently compared by Tukey’s test or Hall-Sidak method (all pairwise multiple comparisons). Differences were considered significant if P0.05. Same letter indicate significantly different from each other. Fig. 1 RESULTS • Animals and in vitro culture of ovarian follicles: Killifish (Fundulus heteroclitus) were collected from salt marshes in St. Augustine, FL. Fish were maintained in a 25 gal. aquarium on a 14/10 hr light/dark cycle, at 25ºC, and were fed three times a day with flake food (TetraMin). • Ovaries were removed from females and placed in 75% Leibovitz L-15 medium with L-glutamine (Sigma) containing 100g gentamicin/ml, pH 7.5. • Fully grown follicles (1.2-1.4 mm in diameter) were manually isolated from several ovaries with the aid of fine forceps under a stereomicroscope, pooled, and randomly distributed into 24-well tissue culture trays (Costar), containing 20 follicles/1ml L-15 media/ well. Incubations were carried out at room temperature and GVBD was scored at 24 and 48 hr. • 17α,20β-dihydroprogesterone (DHP) was obtained from Steraloids Inc.(Newport, RI) and dissolved in ethanol. Porcine inhibin was obtained from Sigma (St. Louis, MO) and dissolved in culture media. Recombinant human activin A was supplied by Dr. Parlow (NIDDK’s National Hormone and Pituitary Program and NICHD) and prepared in culture media. • RNA isolation: mRNA was isolated from Fundulus heteroclitus ovaries (1-2 g) after homogenization of the tissue in a Lysis/binding Solution using the Poly(A)Pure mRNA isolation kit (Ambion). This RNA preparation was used for the reverse transcription reactions (RT) and rapid amplification of 3' and 5' cDNA ends (RACE). Fig. 3 Effects of Activin on DHP-induced Oocyte Maturation CONCLUSIONS • This study provides evidence for the presence of the activin/inhibin ßB subunit in the ovary of F. heteroclitus. • A multiple sequence alignment comparison of the Fundulus Activin/Inhibin Beta B protein sequence with human (gi|9257225| ), chicken (gi|1429377), Xenopus (gi|386028), goldfish (gi|2209351) and zebrafish (gi|516357) sequences (Fig. 6) shows high amino acid sequence homology at the carboxy terminal. Fundulus ßB subunit has well conserved Cysteine positions when aligned with other species. • Results indicate that these gonadal peptides play a paracrine role (acting on neighboring cells) in modulating the oocyte maturation induced by steroid in this species. • The effects of activin and inhibin are distinct and opposed to each other. These effects are reversible, and dependent not only on the concentration of the peptides but also on the dose of the inducing steroid. While inhibin appears to delay the onset of GVBD triggered by the steroid, activin enhances the steroid-induced GVBD by increasing the number of oocytes responding to a low dose of steroid. Recombinant human activin A alone did not induce oocyte maturation (GVBD) in F. heteroclitus. However, DHP-induced GVBD was significantly enhanced by activin A in a dose-dependent manner after 48 hr. No activin effect was observed during the first 24 hr of culture. REFERENCES ACKNOWLEDGEMENTS Sister John Karen Frei, O.P., PhD. Dr. Flona Redway NIH-MBRS SCORE Grant: GM 45455-08 RISE Grant: GM 59244-01A1 NSF Grant: DBI-0116080 Ackland JF, Schwartz NB, Mayo KE, Dodson RE. 1992. Non-steroidal signals originating in the gonads. Physiol Rev 72: 731-787. Alak BM, Smith GD, Woodruff TK, Stouffer RL, Wolf DP. 1996. Enhancement of primate oocyte maturation and fertilization in vitro by inhibin A and activin A. Fertil Steril 66: 646-653. Findlay JK, Hutchinson LA, Risbridger GP. 1987. Intragonadal regulation by inhibin-regulated peptides. In: Burger HG, de Krester DM, Findlay JK, Igarashi M, editors. Inhibin-non-steroidal regulation of follicle stimulating hormone secretion. New York: Raven Press. 42: 299-312. Lin Y-W P, Petrino TR, Landin AM, Franco S, Simeus I. 1999. Inhibitory action of the gonadopeptide inhibin on amphibian (Rana pipiens) steroidogenesis and oocyte maturation. J Exp Zool 284: 232-240. Mather JP, Moore A, Li RH. 1997. Activins, inhibins, and follistatins: further thoughts on a growing family of regulators. Proc Soc Exp Biol Med 215: 209-222. Pang Y, Ge W. 2002. Gonadotropin and activin enhance maturational competence of oocytes in the zebrafish (Danio rerio). Biol Reprod 66: 259-265. Petrino TR, Lin Y-W P, Netherton JC, Powell DH, Wallace RA. 1993. Steroidogenesis in Fundulus heteroclitus V.: purification, characterization and metabolism of 17α,20β-dihydroxy-4-pregnen-3-one by intact follicles and its role in oocyte maturation. Gen Comp Endocrinol 92: 1-15. Stock AE, Woodruff TK, Smith LC. 1997. Effects of inhibin A and activin A during in vitro maturation of bovine oocytes in hormone-and serum-free medium. Biol Reprod 56: 1559-1564. Ying S-Y. 1988. Inhibins, activins, and follistatins: gonadal proteins modulating the secretion of follicle-stimulating hormone. Endocr Rev 9: 267-293.





















