Download

1 / 52
540 likes | 883 Views
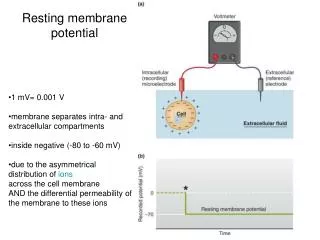
Resting Membrane Potentials. The Lipid Bilayer is a Capacitor. The electric signals of neurons arise from the movement of charges – in the form of ions across the plasma membrane. The membrane has 2 essential features:

E N D
The Lipid Bilayer is a Capacitor • The electric signals of neurons arise from the movement of charges – in the form of ions across the plasma membrane. • The membrane has 2 essential features: 1. The lipid bilayer is an impenetrable barrier to the movement of ions across it. Ions can be stored within the membrane within the “leaky” channels, thereby actually occupying space among the phospholipid tails. Thus, charge can be stored – like an electrical capacitor! That’s because the membrane is leaky to ions, depending on the number and type of leaky channels, and this must be constantly corrected at the expense of ATP.
The lipid bilayer has high charge storage capacity because it is very thin, enabling the stored charge to get VERY close to the corresponding, but opposite charge, lowering their potential energy. • This separation of charges generates an electric field or potential difference across the membrane, given by V = Q/C or charge/capacitance. • Your typical membrane has a capacitance of 1 μF/cm2, about 70% of which is due to the lipid bilayer and the rest because of embedded proteins.
At Rest, the Neuron Membrane is Permeable to K+, Na+ and Cl- Evidence from radioactive tracer studies reveals that all major ionic species can pass through the resting neuronal membrane; i.e., there are “leak channels” for these ions – Outside K+ Na+ Cl- Inside K+Na+ Cl-
However, not all ionic species cross the membrane at the same rate or to the same extent • Not all channels allow the same number of ions to pass per unit time (i.e. carry the same current), maybe due to structural differences in the channels? e.g1. The more narrow the channel the more difficult it is for an ion to pass through the channel (bumps into the wall, squeezes through, resulting in decrease in the kinetic energy) e.g2. There are different numbers of the different channels in the membrane (i.e. there are more K+ leak channels than Na+ leak channels)
Permeability A measure of how easy it is for an ion to cross the membrane; takes into account ease to traverse a channel or the number of channels, etc. At rest, for giant squid axon – Pk+ : PNa+ : PCl- = 1 : 0.04 : 0.45 Ratios differ for different species and different cell types within a species. Here, PK+ is 25 x higher than PNa+. In other neuronal types, in rat, this difference can be as high as 75 x greater.
The Basis of the Resting Membrane Potential [Marieb, E. Human Anatomy and Physiology, 5th ed.]
The Na+-K+ATPase Coupled Pumps to Counters the Resting Ionic Fluxes • At rest, the low level influx of Na+ is balanced by the low level efflux of K+. • This leak must be countered over the long term by the ATP-coupled pumps that maintain the ionic concentration gradients across the membrane.
The ATP-coupled Pump is Electrogenic • At rest, the passive ion fluxes (due to concentration gradients and Vm) are countered by the active ionic fluxes (due to the ATP-coupled pumps). • Consequently, Vm is constant at -60 mV and the K+ and Na+ concentration gradients are also constant through time. • Since the pump moves more Na+ out of the cell than it moves K+ into the cell, it contributes to the Vm, i.e. it contributes to the net negativity inside the cell.
Membrane potentials (Vm) = a voltage difference between the intracellular and extracellular fluids. • Vm is always negative inside relative to outside. • Vm ranges from -40 to -200 mV, depending on the type of cell. • Range of resting Vms for mammalian neurons is -40 to -75 mV; • Any electrical signaling involves deviating from the resting value.
This equilibrium point arises when the ratio of the P(finding an ion on the high-energy side (outside)) P(finding an ion on the low-energy side (inside)) = e-ΔE/kT (i.e., Boltzmann Distribution). The ΔE for the ion is given by Q xVm and because concentrations α probabilities, the result is the Nernst Equation: Vm = EK = (kT/ze)* ln{[K]o/[K]i} Where e is the electronic charge and EK is the equilibrium Nernst potential for K. This can also be written as Ek = 58*log{[K]o/[K]i} More on this later…
Vm results from the asymmetric distribution of charges on either side of the plasma membrane (more negative inside than outside). • However, most of the intracellular and extracellular “fluid” are homogenously distributed (mixed, if you will) on either side of the membrane, making the bulk of the fluid electrically neutral. • Charge separation is achieved in small pockets or clouds (~ 2 μm) of charges spread over the surface and attract each other on either side of the membrane.
For typical cellular values of [K]o & [K]I, EK+ ~ -90 mV, which would be Vm, if the membrane were permeable to only K+. • IMPORTANT! Only a very small fraction – only ~ 1ppm (10-6) have to leave the cell for Vm to reach EK+ .
Electric Charges can arise within Membranes in 2 ways: • Experimenters insert microelectrodes and inject them. • Ion channels are open in the membrane. e.g., K+ is the main ion that establishes the resting membrane potential. This channel is made up of 4 subunits embedded within the membrane
Crystal Structure of the K+ Channel from above and from the side The K+ channel is structure such that a very narrow tube through the inverted cone shape allows for only 50 H2O molecules and only 2 K+ in succession. Because they strongly repel each other, when one enters, one will be forced out.
Two things determine the Voltage across the Membrane • Selective passage of ions through ion channels. • [Ion]s may differ on either side of the membrane. The membrane potential Vm tends to oppose (further) diffusion of K+ since a negative Vm pulls K+ back into the cell. There is a balance between the disordering effect of concentration and the ordering effect of Vm. What is this balance…?
There is also differential distributions of different species of ions (biological charge carriers) across the plasma membrane. E.g.,– giant squid axon (Loligo) Ion [Cytoplasm] [Extracellular Fluid] (mM) (mM) K+ 400 20 Na+ 50 440 Cl- 52 560 Organic- 385 0
Generalizations (although absolute values differ across species and among cells from the same organism) [K+]I (inside) > [K+]o (outside) [Na+]I < [Na+]o [Cl-]I < [Cl-]o [Organic-]I > [Organic-]o
Recall this slide? Resting Membrane Potential, Vm [Marieb, E. Human Anatomy and Physiology, 5th ed.]
Thus, the unique permeability characteristics of the plasma membrane coupled with the original K+ concentration gradient leads to the establishment and maintenance of the Vm without expenditure of energy.
Use of the Nernst Equation to Calculate Theoretical Vm for a Membrane Permeable Only to K+ E = RT/zFln [ion]o(outside)/[ion]I(inside) E = membrane potential (Volts) R = gas constant (8.3143 joules/deg-mole) T = absolute temperature (273o + oC); usually 20oC for giant squid axon z = valence, including charge and number F = Faraday’s constant (96,490 coulombs/mole)
Simplifying the Equation • RT/zF reduces to 0.025 Volts or 25 mVolts at 20oC and a valence of +1 (K+ and Na+) • Thus, E (mVolts) = 25 ln [ion]o/[ion]I • Converting to log10 : E (mV) = 25 x 2.3 log [ion]o/[ion]I E (mV) = 58 log [ion]o/[ion]I
If the Nernst Equation is a Good Model of Membrane Potential Development, What Should Happen if We Change [K+]o? Predict Vm when [K+]o = [K+]I
Semi log plot of Vm versus [K+]outside, i.e. changing [K+]o changes the concentration gradient across the membrane and the Vm that develops in response to that gradient 0 Vm (mV)-60 -120 Normal Vm of -75 mV Normal [K+]o 4 20 400 Log [K+]outside
This is a common experimental technique used when you want to stimulate (excite) (a) neuron(s), say, in tissue culture or in brain slices. • The addition of KCl to the bath or medium will result in a depolarization – (Why?). • However, as we will see when we cover action potentials, the preceding slide is applicable only up to T0, given the all-or-none nature of the AP.
What About Cl-? 1) Cl- permeability is relatively high 2) However, there is no active mechanism to move Cl- across the membrane of most cells. 3) Consequently, Cl- passively distributes itself across the membrane in relation to the Vm established by Na+ and K+.
Flipped because of negative valence on Cl- Vresting = 58 log [Cl-]I/[Cl-]o Set by Na+ and K+ Cl- passively distributes itself across the membrane such that the concentration gradient balances the Vm. Net - Net + Concentration gradient Cl- Cl- Electrical gradient = Vm
A Modification of the Nernst Equation is the Goldman-Hodgkin Equation can be used to Predict Vm when the Membrane is Permeable to Multiple Ions Vmtheoretical = - 60 mV ~ the empirically measured value in a resting neuron
What happens to the Vm when leak channels for Na+ and Cl- are introduced into the membrane? Exercise #1 = Draw equilibrium in a resting neuron. Start with a liver cell in equilibriumand add Na+ leak channels to the membrane. (Hint #1: See Time0+1 when Na+ is just starting to enter the cell - the second image to follow this slide). (Hint #2 : at equilibrium Vm = -60 mV and [K+]I>[K+]O and [Na+]I<[Na+]O
Explanation • Na+ enters the cell and depolarizes the membrane. The rate of entry is low and is set by the # and structure of the Na+ leak channels and the Na+ concentration gradient. • The Vm no longer balances the K+ concentration gradient. • A small amount of K+ is now able to leave the cell. • At equilibrium, the rate of entry of Na+ is equal to the rate of exit of K+ and the Vm is constant at a new value of -60 mV; i.e. more + than for “liver cell” equilibrium. At equilibrium there is no net movement of charge.
Explanation (cont’d) 5) The Vresting is much closer to EK+ than ENa+ because the permeability of the membrane to K+ is much greater than the permeability of the membrane to Na+.
A Simple Neural Network to Demonstrate Signaling within the Nervous System Synapse = site of cell to cell communication in the nervous system; electrical signal gets converted to a chemical signal (neurotransmitter) then back into an electrical signal Presynaptic neuron = carries information as an electrical signal (action potential) toward the synapse Postsynaptic neuron = carries information as an electrical signal away from the synapse
Membrane Potentials - to recap: A. All living cells have membrane potentials (Vm) = a voltage difference between the intracellular and extracellular fluids; they are always negative inside relative to outside (convention in neurophysiology); Vm ranges from -40 to -200 mV(olts) depending on the type of cell; mammalian neurons have a range of resting Vms from -40 to -75 mV and electrical signaling involves a change away from the resting value
1) Recording a Vm– begin with both electrodes in the extracellular fluid Recording apparatus = ohmmeter, amplifier, oscilloscope or computer Intracellular electrode = glass pipette drawn out to a small tip diameter (~0.5 um); filled with highly conductive KCl solution AgCl electrodes (wires) KCl KCl Liver cell = non-excitable cell Extracelluar electrode = larger glass pipette also filled with KCl solution
+75 Vm (mVolts)0 -75 1 2 Time (sec) KCl Liver cell = non-excitable cell KCl 2) Recording a Vm– impale cell with intracellular electrode
B. Membrane Potentials Result from Differential Distributions of Electrical Charges Across the Plasma Membrane; but the bulk of the intracellular and extracellular fluids are electrically neutral; charge separation exists in a small cloud of ~2 um spread over the intracellular and extracellular surfaces of the plasma membrane ++++++++++++++++++++++++++++++++++++++++++++ ++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ ++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ +++++++++++ - - - - - - - - - - - ++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++++ - - - + + + - - - + + + - - - - - - - - - - +++++++++++ ++++++++++++++++++++++++++++++++++++++++++++
Remember this slide from earlier? There is a differential distributions of different species of ions (biological charge carriers) across the plasma membrane. E.g.,– giant squid axon (Loligo) Ion [Cytoplasm] [Extracellular Fluid] (mM) (mM) K+ 400 20 Na+ 50 440 Cl- 52 560 Organic- 385 0
And this? Generalizations (although absolute values differ across species and among cells from the same organism) [K+]I (inside) > [K+]o (outside) [Na+]I < [Na+]o [Cl-]I < [Cl-]o [Organic-]I > [Organic-]o
Building a Model of Membrane Function that Explains Empirical Measures of Vm and Ionic Concentration Differences Exercise #1 = draw equilibrium for a membrane freely permeable to all cations, i.e. the laws of simple diffusion explain the observations. (See following slide.) Hint = Vm is 0 mV at time0 and timeeq
Time0 Vm = 0 mV K+ Na+ K+ Na+ “chemical force” magnitude and direction of net diffusion Hypothetical membrane freely permeable to all cations inside outside
Timeeq Vm = 0 mV K+ Na+ K+ Na+ “chemical force” magnitude and direction of net diffusion • Explain: • Equal sizes of ions • Double-headed arrows • No Vm Hypothetical membrane freely permeable to all cations inside outside
Answers:1) Diffusion continues until both cations are equally distributed across the membrane (law of diffusion).2) The membrane is freely permeable to both cations so they continue to move across the membrane once equilibrium is achieved, but there is no NET exchange of the ions across the membrane.3) The original cation concentration gradients were the same approximate magnitude, so when ionic concentrations equilibrate across the membrane the same number of positive charges have left the cell as have moved into the cell. Therefore no charge separation, i.e. Vm, has been created.
What do we do with this model of membrane function? Reject it because it does not agree with empirical measures of Vm and ionic concentration gradients
Building a Model of Membrane Function that Explains Empirical Measures of Vm and Ionic Concentration Differences Exercise #2 = draw Time0+3xK+ exit for a membrane permeable only to K+. (See following slide.) Hint = Vm is 0 mV at time0
= Membrane is impermeable to ion Time0 Vm = 0 mV K+ Na+ Cl- Organic- K+ Na+ Cl- Organic- “chemical force” magnitude and direction of net diffusion Hypothetical membrane freely permeable to only K+ inside outside
Time0+3xK+ exit Vm = developing = Membrane is impermeable to ion K+ Na+ Cl- Organic- K+ Na+ Cl- Organic- - - - + + + “chemical force” magnitude and direction of net diffusion “electrical force” (and direction of movement) developing Hypothetical membrane freely permeable to only K+ inside outside
Answers:1) Diffusion of K+out of the cell begins at Time0 and at Time0+3xK+ exit 3 + charges (in the form of potassium) have left the cell, leaving net negativity behind inside the cell. 2) The + charges outside the cell repel other + charges (or the – charges inside the cell attract + charges) causing some K+ to move back into the cell. This charge separation is nothing more than a Vm developing across the plasma membrane.3) At this point, the “chemical force” is still greater than the “electrical force” and there is net movement of K+ out of the cell.
Building a Model of Membrane Function that Explains Empirical Measures of Vm and Ionic Concentration Differences Exercise #2 = draw Timeeqfor a membrane permeable only to K+. Hint = at Timeeq there is no NET exchange of K+ across the membrane and [K+]I > [K+]o
Timeeq Vm = -75 mV = Membrane is impermeable to ion K+ Na+ Cl- Organic- K+ Na+ Cl- Organic- - - - - - - - - - “chemical force” magnitude and direction of net diffusion + + + + + + + + + “electrical force” = - “chemical force” Hypothetical membrane freely permeable to only K+ inside outside