Download
1 / 1
10 likes | 106 Views
Experiment 1. Fig. 1. Fig. 2. Fig. 6. Fig. 5. Fig. 7. Fig. 8. Fig. 3. Fig. 4. Fig. 9. Fig. 10. Fitness Problems in Escherichia coli K-12 Transformed with a High Copy Plasmid Encoding the Green Fluorescent Protein

E N D
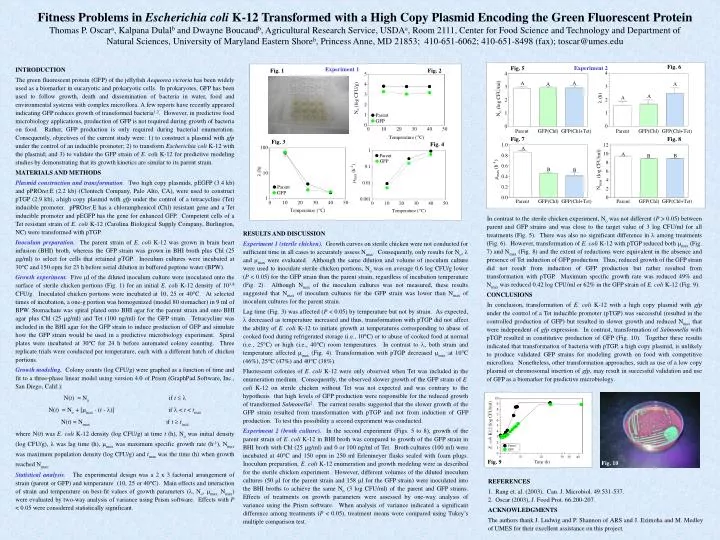
Experiment 1 Fig. 1 Fig. 2 Fig. 6 Fig. 5 Fig. 7 Fig. 8 Fig. 3 Fig. 4 Fig. 9 Fig. 10 Fitness Problems in Escherichia coli K-12 Transformed with a High Copy Plasmid Encoding the Green Fluorescent Protein Thomas P. Oscara, Kalpana Dulalb and Dwayne Boucaudb, Agricultural Research Service, USDAa, Room 2111, Center for Food Science and Technology and Department of Natural Sciences, University of Maryland Eastern Shoreb, Princess Anne, MD 21853; 410-651-6062; 410-651-8498 (fax); toscar@umes.edu Experiment 2 INTRODUCTION The green fluorescent protein (GFP) of the jellyfish Aequorea victoria has been widely used as a biomarker in eucaryotic and prokaryotic cells. In prokaryotes, GFP has been used to follow growth, death and dissemination of bacteria in water, food and environmental systems with complex microflora. A few reports have recently appeared indicating GFP reduces growth of transformed bacteria1,2. However, in predictive food microbiology applications, production of GFP is not required during growth of bacteria on food. Rather, GFP production is only required during bacterial enumeration. Consequently, objectives of the current study were: 1) to construct a plasmid with gfp under the control of an inducible promoter; 2) to transform Escherichia coli K-12 with the plasmid; and 3) to validate the GFP strain of E. coli K-12 for predictive modeling studies by demonstrating that its growth kinetics are similar to its parent strain. MATERIALS AND METHODS Plasmid construction and transformation. Two high copy plasmids, pEGFP (3.4 kb) and pPROtet.E (2.2 kb) (Clontech Company, Palo Alto, CA), were used to construct pTGP (2.9 kb), ahigh copy plasmid with gfp under the control of a tetracycline (Tet) inducible promoter. pPROtet.E has a chloramphenicol (Chl) resistant gene and a Tet inducible promoter and pEGFP has the gene for enhanced GFP. Competent cells of a Tetresistant strain of E. coli K-12 (Carolina Biological Supply Company, Burlington, NC) were transformed with pTGP. Inoculum preparation. The parent strain of E. coli K-12 was grown in brain heart infusion (BHI) broth, whereas the GFP strain was grown in BHI broth plus Chl (25 g/ml) to select for cells that retained pTGP. Inoculum cultures were incubated at 30C and 150 opm for 23 h before serial dilution in buffered peptone water (BPW). Growth experiment. Five l of the diluted inoculum culture were inoculated onto the surface of sterile chicken portions (Fig. 1) for an initial E. coli K-12 density of 103.8 CFU/g. Inoculated chicken portions were incubated at 10, 25 or 40C. At selected times of incubation, a one-g portion was homogenized (model 80 stomacher) in 9 ml of BPW. Stomachate was spiral plated onto BHI agar for the parent strain and onto BHI agar plus Chl (25 g/ml) and Tet (100 ng/ml) for the GFP strain. Tetracycline was included in the BHI agar for the GFP strain to induce production of GFP and simulate how the GFP strain would be used in a predictive microbiology experiment. Spiral plates were incubated at 30C for 24 h before automated colony counting. Three replicate trials were conducted per temperature, each with a different batch of chicken portions. Growth modeling.Colony counts (log CFU/g) were graphed as a function of time and fit to a three-phase linear model using version 4.0 of Prism (GraphPad Software, Inc., San Diego, Calif.): N(t) = No if t N(t) = No + [max (t - )] if < t < tmax N(t) = Nmax if ttmax where N(t) was E. coli K-12 density (log CFU/g) at time t (h), No was initial density (log CFU/g), was lag time (h), max was maximum specific growth rate (h-1), Nmax was maximum population density (log CFU/g) and tmax was the time (h) when growth reached Nmax. Statistical analysis. The experimental design was a 2 x 3 factorial arrangement of strain (parent or GFP) and temperature (10, 25 or 40C). Main effects and interaction of strain and temperature on best-fit values of growth parameters (, No, max, Nmax) were evaluated by two-way analysis of variance using Prism software. Effects with P < 0.05 were considered statistically significant. In contrast to the sterile chicken experiment, No was not different (P > 0.05) between parent and GFP strains and was close to the target value of 3 log CFU/ml for all treatments (Fig. 5). There was also no significant difference in among treatments (Fig. 6). However, transformation of E. coli K-12 with pTGP reduced both max (Fig. 7) and Nmax (Fig. 8) and the extent of reductions were equivalent in the absence and presence of Tet induction of GFP production. Thus, reduced growth of the GFP strain did not result from induction of GFP production but rather resulted from transformation with pTGP. Maximum specific growth rate was reduced 49% and Nmax was reduced 0.42 log CFU/ml or 62% in the GFP strain of E. coli K-12 (Fig. 9). CONCLUSIONS In conclusion, transformation of E. coli K-12 with a high copy plasmid with gfp under the control of a Tet inducible promoter (pTGP) was successful (resulted in the controlled production of GFP) but resulted in slower growth and reduced Nmax that were independent of gfp expression. In contrast, transformation of Salmonella with pTGP resulted in constitutive production of GFP (Fig. 10). Together these results indicated that transformation of bacteria with pTGP, a high copy plasmid, is unlikely to produce validated GFP strains for modeling growth on food with competitive microflora. Nonetheless, other transformation approaches, such as use of a low copy plasmid or chromosomal insertion of gfp, may result in successful validation and use of GFP as a biomarker for predictive microbiology. RESULTS AND DISCUSSION Experiment 1 (sterile chicken). Growth curves on sterile chicken were not conducted for sufficient time in all cases to accurately assess Nmax. Consequently, only results for No, and max were evaluated. Although the same dilution and volume of inoculum culture were used to inoculate sterile chicken portions, No was on average 0.6 log CFU/g lower (P < 0.05) for the GFP strain than the parent strain, regardless of incubation temperature (Fig. 2). Although Nmax of the inoculum cultures was not measured, these results suggested that Nmax of inoculum cultures for the GFP strain was lower than Nmax of inoculum cultures for the parent strain. Lag time (Fig. 3) was affected (P < 0.05) by temperature but not by strain. As expected, decreased as temperature increased and thus, transformation with pTGP did not affect the ability of E. coli K-12 to initiate growth at temperatures corresponding to abuse of cooked food during refrigerated storage (i.e., 10C) or to abuse of cooked food at normal (i.e., 25C) or high (i.e., 40C) room temperatures. In contrast to , both strain and temperature affected max (Fig. 4). Transformation with pTGP decreased max at 10C (46%), 25C (47%) and 40C (38%). Fluorescent colonies of E. coli K-12 were only observed when Tet was included in the enumeration medium. Consequently, the observed slower growth of the GFP strain of E. coli K-12 on sterile chicken without Tet was not expected and was contrary to the hypothesis that high levels of GFP production were responsible for the reduced growth of transformed Salmonella2. The current results suggested that the slower growth of the GFP strain resulted from transformation with pTGP and not from induction of GFP production. To test this possibility a second experiment was conducted. Experiment 2 (broth culture). In the second experiment (Figs. 5 to 8), growth of the parent strain of E. coli K-12 in BHI broth was compared to growth of the GFP strain in BHI broth with Chl (25 g/ml) and 0 or 100 ng/ml of Tet. Broth cultures (100 ml) were incubated at 40C and 150 opm in 250 ml Erlenmeyer flasks sealed with foam plugs. Inoculum preparation, E. coli K-12enumeration and growth modeling were as described for the sterile chicken experiment. However, different volumes of the diluted inoculum cultures (50 l for the parent strain and 158 l for the GFP strain) were inoculated into the BHI broths to achieve the same No (3 log CFU/ml) of the parent and GFP strains. Effects of treatments on growth parameters were assessed by one-way analysis of variance using the Prism software. When analysis of variance indicated a significant difference among treatments (P < 0.05), treatment means were compared using Tukey’s multiple comparison test. • REFERENCES • Rang et. al. (2003), Can. J. Microbiol. 49:531-537. • Oscar (2003), J. Food Prot. 66:200-207. • ACKNOWLEDGMENTS • The authors thank J. Ludwig and P. Shannon of ARS and J. Ezimoha and M. Medley of UMES for their excellent assistance on this project.






