Download

1 / 71
730 likes | 905 Views
Signal Transduction. First lecture. Advanced Biochemistry Course. March 2009. Plasma Membrane Structure and Function. The plasma membrane separates the internal environment of the cell from its surroundings The plasma membrane is a phospholipid bilayer with embedded proteins .

E N D
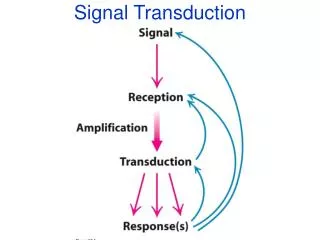
Signal Transduction First lecture Advanced Biochemistry Course March 2009
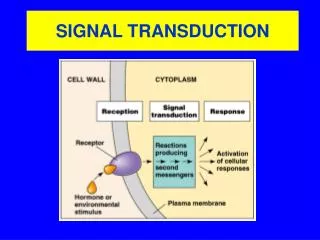
Plasma Membrane Structure and Function The plasma membraneseparates the internal environment of the cell from its surroundings The plasma membrane is aphospholipid bilayerwithembedded proteins. The plasma membrane has afluidconsistency and amosaicpattern of embedded proteins.
Protein dynamics in the lipid bilayer 1) Proteins can move laterally in the plane of the membrane (capping) 2) Proteins can rotate around an axis vertical to the plan of the membrane (channels) 3) Proteins cannot tumble through the plan of the membrane
Lateral Rotational Thumbling
Signal Transduction Endocrinic- “ (Insulin; adrenalin) Paracrinic -transduction (histamin;prostaglandins) Autocrinic - “ (TGF/ IGF) Synaptic - transmission (neurotransmitters)
Receptors Membrane receptors Cytosolic receptors G-coupled receptors Channel/receptors 3. Enzyme/receptors Steroid receptors Vitamin D 3. Retinoic acid
Gramicidin A in lipid bilayer and water • Antibiotic peptide Forms a pore in the cell wall of a bacteria and lets out monovalent cations (K+, Na+). [Membrane potential disappears and bacteria dies!] • 15 amino acids, helical • Channel is formed by a head- to-head dimer
3.5 nm 3.5 nm 3.5 nm 4 nm Glycophorin
Hydropathy plot The Free Energy for Transferring a Helix of 20 Residues from the Membrane to Water
Membrane topology Rhodopsin b-adrenergic receptor
G-protein-coupled receptors definition • One of the largest families of membrane proteins • Common structural architecture • Extracellular N-terminal domain Glycosylated Ligand recognition • Intracellular C-terminal domain Contains several putative phosphorylation sites Involved in desensitisation/internalisation • 7 membrane spanning domains • Couple to G-proteins signal transduction • Divided into subfamilies based on sequence homology
Orphan GPCRs How many GPCRs are encoded by the human genome? • First estimation based on comparison with C. elegans ~1000 GPCRs ~5% of genome 19.100 genes 27.000 genes HuGo ~1800 GPCRs • Current estimation ~1000 GPCRs in human genome ~700 olfactory, gustatory and chemokinine receptors ~300-400 transmitter GPCRs
Orphan GPCRs 210 GPCRs bind un known natural ligands ~400 transmitter GPCRs 160 orphan GPCRs remain to be characterised
B. GPCRs in disease states • Disease states associated with GPCR mutations • Rhodopsin receptor retinitis pigmentosa • Vasopressin V2 nephrogenic diabetes • Glucagon diabetes, hypertension Rationale for oGPCR characterisation Why is the pharmaceutical industry interested in oGPCRs? A. GPCRs are good drug targets • 50% of subscription drugs interact with GPCRs • Hypertension • Stomach ulcers • Migraine • Allergies GRF-receptor dwarfism Asp(60) Gly (60)
GPCR subfamilies Family A: Rhodopsine-like • Largest family • Conserved DRY motif (i2) • Conserved cysteines -S-S- Family B: Secretine-like • Large N-terminal domain • Several well conserved cysteine residues • High Mr hormone ligands Family C: Metabotropic glutamate • Long N-terminal domain • N-terminus sufficient for ligand binding
Protease activated receptor (PAR) R R* S-F-L-L-R-N
The positioning of the the 7TMR in the membrane Lipid head groups Chains hydrophobic Lipid head groups Polar/negatively charged
G-protein-coupled receptors Extracellular -NH2 e3 e2 -S-S- e1 TM1 TM2 TM3 TM4 TM5 TM6 TM7 D R Y i2 i1 i3 Cytoplasmic COOH-
How does the ligand activates the receptor in a selective way? A two state model is commonly used to characterize this activation Fluorescence spectroscopy was used to characterize the diversity of conformational states of the 2AR and its mode of activation
AGONIST-INVERSE AGONIST - ANTAGONIST • Drug effects can be classified into three major phenotypes: agonist, antagonistand inverse agonist. • Agonist and inverse agonisteffects are associated with receptor activation and inactivation, respectively • Antagonismimplies that a drug produces no effect when administered alone but blocks the effects of agonists and inverse agonists.
Agonist- a ligand that activates the receptor Antagonist- a ligand that inhibits the receptor Partial agonist- a low affinity agonist Inverse agonist - a ligand that prefers the inactive form of the receptor Two- State Model r R* Inactive form Active form
Receptor states and inverse agonists (Two-State Model) Partial agonists and antagonists bind to both r and R states rR inactive Inverse Agonist active Agonist rA R*A Activation in the absence of an agonist; over-expression
Energy landscape diagram describing a possible mechanism of GPCR activation by an agonist
1)Inverse agonist (propranolol) binds to the r form of the receptor and the activity of the system is suppressed below its normal spontaneous state2) In between full and inverse agonists are those agonists that bind to both r and R states. These are partial agonists. These are unable to achieve maximal stimulation even if all receptor binding sites are occupiedInverse agonism offers a potential of developing new drugs that attenuate the effect of mutant receptors that are constitutively active3) The neutral antagonists (b-blockers such as pindolol) bind to both r and R conformations and are better regarded as passive antagonistsThey impede the binding of both agonists and inverse agonists.Therefore, pindolol affects the heart only during exercise and stress while propranolol also suppresses the resting heart rate
Activation of G-protein-coupled receptors Ligand Efficacy: The effect of different classes of drugs on a GPCR that has some detectable basal activity
FULL AGONIST PATRIAL AGONIST ANTAGONIST INVERSE AGONIST
Rhodopsin Dopamine
How can we determine the mechanism of activation? Three different methods are used, which could be applied to explore the mechanism of activation of 7TMR or any other receptors The focus will be on the adrenergic receptors
F R E T fluorescence resonance energy transfer A donor chromophore, in its electronic excited state, may transfer energy to an acceptor chromophore (in close proximity < 10nm) through Non-radiative dipole-dipole coupling When both chromophores are fluorescent, the term "fluorescence resonance energy transfer (FRET)" is often used instead, although the energy is not actually transferred by fluorescence
Fluorescence resonance energy transfer (FRET) The two fluorescent probes report in real time through a fast decrease in FRET the intra-molecular conformational rearrangements associated with receptor activation
Agonist Norepinephrine (NE) Inverse agonist Yohimbine
Binding site for norepinephrine in the b2AR Swaminath, G. et al. J. Biol. Chem. 2004;279:686-691
Top-down view of hormone receptor with an adrenalin molecule
weak partial agonist good partial Agonist selective Agonist selective Agonist selective Agonist inverse Agonist antagonist
Arrangement of transmembrane domains of a prototypical G protein-coupled receptor as viewed from the extracellular surface of the membrane (based on the projection maps from two-dimensional crystals of rhodopsin) The Asp3-Arg3 pair at the cytoplasmic end of transmembrane domain 3 (TM3) is part of the highly conserved (D/E)RY motif found in beta2-AR and other rhodopsin-family GPCRs, whereas the Glu6at the cytoplasmic end of TM6 is highly conserved in amine and opsin receptors. The ionic link between the Asp3-Arg3pair and Glu6 is known as the ionic lock Gether et al., J. Biol. Chem. 1998;273:17979
Break ionic lock Activate rotamer toggle switch
The Asp3-Arg3 (D/R) pair at the cytoplasmic end of transmembrane domain 3 (TM3) is part of the highly conserved (D/E)RY motif found in b2AR and other rhodopsin-family GPCRs, whereas the Glu6 at the cytoplasmic end of TM6 is highly conserved in amine and opsin receptors. The ionic link between the Asp3-Arg3 pair and Glu6 is known as the ionic lock The Ionic Link
How is the receptor activated ? • Previous biophysical studies on the b2-AR suggest that agonist binding and activation occurs through at least one conformational intermediate, implying that at least one molecular switch is involved. • These studies also show that structurally different agonists and partial agonists differ in their ability to induce specific conformational transitions.
Cellular responses to catecholamines cAMP accumulation in HEK cells expressing 2AR The state that stabilizes the binding of the catechol ring alone (R1) is not sufficient to activate the G proteins Swaminath, G. et al. J. Biol. Chem. 2004;279:686-691
Fluorescence spectroscopy to monitor disruption of the ionic lock at the b AR
Fluorescence spectroscopy to monitor disruption of the ionic lock at the b AR The bimane-tryptophan technique Mutated A271C and binding of mBrBimane Mutated I135W Relies on the quenching of of bimane fluorescence by Trp at near contact distance in the 5-15 Å range
Quenching during agonist binding to the site
TM6 E 268 TM3 D R Y Asp-Arg-Tyr TM 6 TM 3 A271C Ile135W
Effect of ligand structure on the ionic lock Iso Epi Norepi Dop % maximal quenching Sal Log[ligand] Hal Cat
E268 R131 D130 Ionic Lock Active Inactive A135W H271C-Bimane • DRY
Molecular three-dimensional representations of the interaction of TM3 and TM6 at their cytoplasmic ends and the effects of 6.30 mutations Ballesteros, J. A. et al. J. Biol. Chem. 2001;276:29171-29177