Download

1 / 107
1.12k likes | 1.46k Views
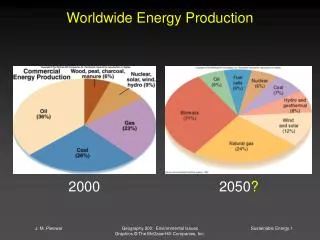
Salinity Impact on Crop Production Worldwide. World Land Surface Area. 150 x 10. km. 6. 2. Salt affected. 9 x 10. km. (6%). 6 . 2 . Cultivated Land. 15 x 10. km. 6 . 2 . *. Salt affected. 2 x 10. km. (13%). 6. 2. Irrigated Land. 2.4 x 10. km. 6. 2. *.

E N D
Salinity Impact on Crop Production Worldwide World Land Surface Area 150 x 10 km 6 2 Salt affected 9 x 10 km (6%) 6 2 Cultivated Land 15 x 10 km 6 2 * Salt affected 2 x 10 km (13%) 6 2 Irrigated Land 2.4 x 10 km 6 2 * Salt affected 1.2 x 10 km (50%) 6 2 *Problem is increasing Negative Impacts of Salinity on Agriculture Reduced yields on land that is presently cultivated Limited expansion into new areas Glycophytes vs halophytes - sweet plants and salt plants, respectively, by definition halophytes are “native flora to a saline environment” Quantitative difference - adaptation Nearly all salt tolerant plants are angiosperms, indicating polyphyletic origin, or halophytes are primitive genetic remnants of different families Salt tolerant species exist in 1/3 of the angiosperm families; however about ½ of the 500 halophytic species belong to 20 families, monocots - 45 genera in the Poaceae family and dicots - 44% of the halophytic genera are in the Chenopodiaceae (Atriplex, Salicornia and Suaeda) Most plants, including the majority of crop species, are glycophytes and cannot tolerate high salinity.
Salt tolerance research is important basic plant biology • Salt tolerance research contributes to our understanding of subjects ranging from gene regulation and signal transduction to ion transport, osmoregulation and mineral nutrition. • Additionally,some aspects of salt stress responses are intimately related to drought and cold stress responses. • Plant salt tolerance studies thus contribute to understanding cross-tolerance
Evolution of salt tolerance Soil salinity almost always originates from previous exposure to seawater Although it is believedthat for most of the Earth's history, the salt level of theoceans was much lower than now,all plant species that inhabit the seas, as well as a phylogeneticallydiverse groups of land plants, are capable of growth and reproductionat salinity levels near or above those found in the seas. This strongly supports the existenceof a genetic basis for high-salinity tolerance within both seaand land plants. Plant Physiol. 135, 1718-1737.
Sensitivity to salt occurs during all plant growth stages germination NaCl inhibits both 1) germination and 2) growth למה?
Resistance to drought and salt stresses by neutrally charged osmolytes Osmolytes Heat shock proteins LEA Compatible solutes protect the hydration shell
Osmolytes/Osmoprotectants. Listed are common osmolytes involved in either osmotic adjustment or in the protection of structure. In all cases, protection has been shown to be associated with accumulation of these metabolites, either in naturally evolved systems or in transgenic plants
Secondary effects of NaCl stress Reduced cell expansion and assimilate production – as during drought, adaptation includes reduction in cell expansion that affects photosynthate production Photosynthate production is reduced – carbon metabolism is salt sensitive Decreased cytosolic metabolism – metabolic poisoning, althoughenzymes of halophytes and glycophytes are equally sensitive to NaCl Production of ROS – products of photorespiration and mitochondrial respiration when electron flow is too great for the normal electron acceptors of metabolism, e.g. NADPH, resulting in the production of ROS A. spongiosa and S. australis are halophytes If the halophytes’ and glycophytes’ enzymes are equally sensitive to NaCl, why are the plants differentially sensitive to NaCl?
Ion disequilibrium – Na+ rapidly enters the cell because the membrane potential inside is negative (~-120 to -200 mV), see slide Signal_transduction_of_Responses_to_Environment.ppt#17. Ionomics Na+ can accumulate to102- to 103-fold greater concentration than in the apoplast, driven by the membrane potential, sea water 457 mM Na+ Na+ is cytotoxic, while K+ is an essential nutrient Ca2+ disequilibrium affects K+/Na+ selective uptake some plant species are also sensitive to Cl- Cultured tobacco (glycophyte) cells are inhibited by 100 mM NaCl; however, after adaptationtobacco cells can grow in 500 mM NaCl Thus, the salt tolerance mechanism exists in glycophytes
Selective ion uptake and differential ion compartmentalization are main features that explain salt tolerance disparity between glycophytes and halophytes • (Flowers et al., 1977; Greenway and Munns, 1980; Jeschke, 1984). Salinity affects nutrient acquisition by interfering with K+ uptake by carriers and channels. At the cellular level, intracellular ion sequestration into vacuoles for osmotic adjustment, strong ion selectivity in the cytosol (preference of K+ over Na+), and accumulation of compatible (non-toxic) organic solutes in the cytosol to equilibrate water potential across the tonoplast are widely accepted mechanisms contributing to salt tolerance (Greenway and Munns, 1980; Gorham et al., 1985). The sequestration of ions that are potentially damaging to cellular metabolism (e.g. Cl–, Na+) into the vacuole while maintaining high K+/Na+ ratios in the cytosol would provide the osmotic driving force required for water uptake in saline environments and, at the same time, provide plants with an efficient instrument for ion detoxification
NaCl induces cytological hallmarks of programmed cell death in the wild-type yeast Nuclear fragmentation also IN PLANTS Bc2-2 protects Nuclear fragmentation (1h) normal mitochondrion abnormal mitochondrion d) Nuclear fragmentation; e) vacuolation; f) coalescence of vacuolar and nuclear membranes; g) cell lysis. Hamilton,E & Heckathorn, S (2001) Plant Physiol. 126, 1266-1274. IN PLANTS Mitochondrial adaptations to NaCl. Complex I is Protected by Anti-Oxidants and Small Heat Shock Proteins, whereas Complex II is Protected by Proline and Betaine.
NaCl Uptake into Roots and Movement in the Plant Radial transport from the soil solution into roots is apoplastic/symplastic (epidermis and cortex), symplastic across the endodermis and then loaded into the xylem Radial transport may be regulated, i.e., Na+ and Cl- transport to the xylem is limited in epidermal and cortical cells, i.e., prior to the endodermis, but xylem loading is passive, plants can regulate K+/Na+ concentration in the xylem sap. Casparian strip ensures that all substances pass through at least one membrane before entering the stele Salt movement through the xylem is determined by the transpirational flux – moves through the xylem to the shoot Plants minimize exposure of meristematic cells to Na+and Cl- - the lack of vasculature to the meristem reduces transport to these cells, mature leaves are ion sinks and may abscise Some halophytes deposit salt on the surface of leaves (sink) via glands or bladders
development of salt-tolerant crops (i.e. accumulation of salt) Twenty years ago, Epstein argued for the development of salt-tolerant crops with truly halophytic responses to salinity in which the consumable part is botanically a fruit, such as grain or berries or pomes. In these plants, Na+ would accumulate mainly in their leaves and, because the water transport to the fruits and seeds is mainly symplastic, much of the salt would be screened from these organs. Thus, engineering theaccumulation of salt in vacuolated cells, together with the active extrusion of Na+ from non-vacuolated cells (i.e. young and meristematic tissue),will allow the maintenance of a high cytosolic K+/ Na+ ratio. In combination with the enhanced production of compatible solutes…
Studying the Salt stress • 1) Physiology of salt toxicity and salt tolerance. This includes cellular and metabolic responses to salt(Bohnert and Sheveleva, 1998 ; Hasagewa et al., 2000 ), as well as whole plant responses (Flowers et al., 1997; Greenway and Munns, 1980 ; Yeo, 1998 ). • 2) Mechanisms of salt transport across cellular membranes and over long distances. This includes physiological and molecular characterization of ion transporters involved in salt uptake, extrusion, compartmentalization(Blumwald et al. 2000 ; Schachtman and Liu, 1999 ). • 3) Survey genes whose expression is regulated by salt stress (Zhu et al., 1997 ; Xiong and Zhu, 2001 ; Shinozaki and Yamaguchi-Shinozaki, 1997 ; Ingram and Bartels, 1996 ; Bray, 1997 ; Bohnert et al., 1995 ). This research is accelerated by using microarrays (Seki et al., 2001 ; Kawasaki et al., 2001 ; Bohnert et al., 2001 ). • 4) Mutational analysis of salt tolerance determinants and salt stress signaling(Zhu, 2000 ; 2001a , b ; Xiong and Zhu, 2001 ).
Salt research approaches I • Comparative biochemistry(between species, treatments) • osmolytes • ROS • ion compartmentation mechanisms(Na+ enters root cells mainly through various cation channels, particularly voltage-(in)dependent cation channels. Na+ and K+ • Mutants (UpOR Down) • Overexpression of individual components • Complementation of yeast mutants inhibitors of salt adaptation in yeast
Functional Genomics of Plant Stress Tolerance • Complexity and Multigenicity of Stress Responses. • 1. Variations on common physiological Themes. • 2.Evolutionary Conservation of Stress Responses Mutants with altered sensitivity to osmotic/salt stress Mutants in stress signal transduction pathways using osmotically regulated promoter-reporter screening Identify Suppressors of Stress-responsive mutants
salt cress Unlike Arabidopsis leaf morphology,salt cress displays succulent-like leaves after salt exposure,measured as FW to dry-weight ratio. The developmentof a second layer of leaf palisade cells maycontribute to this and also affect the rate of water loss fromleaves. In addition, the stomatal density on salt cress leavesis twice that of Arabidopsis, althoughthe stomatal index is nearly the same This may allow more efficient distribution ofCO2 to photosynthetic mesophyll cells at low stomatal apertures.Plant Physiol.135, 1718 The difference in salt sensitivity/tolerance may have resulted from differences in regulatory circuits or from salt tolerance genes. For example, the vacuolar Na+/H+ antiporter gene AtNHX1 is not as highly inducible in Arabidopsis as its homologous gene is in halophytes, and high level AtNHX1 expression driven by the strong CaMV 35S promoter could significantly improve Arabidopsis salt tolerance (Apse et al., 1999 ; Hamada et al., 2001 ; Shi and Zhu, 2002 ). At Book
The role of Potassium (K+) Potassium affects the life of every living being. K+ role in plant growth is quite similar to that for humans. K+ is not an integral part of organic molecules in plants. K+ is important in many biochemical reactions, e.g. translocation of carbohydrates Under severe deficiency, plants will often develop visible symptoms: older leaf edges will turn brown, yield and quality decline. Sometimes, orange trees will drop their fruit; strawberries do not develop their sweet taste; corn stalks will break; tomatoes will be small and contain too much white tissue. Alfalfa will show typical yellowing along the outer margins of the leaves.
Salt stress impairs K nutrition The membrane potential difference at the plasma membrane of plant cells is -140 mV, which favors passive transport of Na+ into cells, especially with high extracellular Na+ concentrations. Excess extracellular Na+ enters the cell through both the transporter HKT1 and non-selective cation channels/ transporters, which results in a decrease in the K+/Na+ ratio in the cytosol. With increased concentration of NaCl in the medium, Na+ ↑ whereas K+↓. Analysis of ion content in seedlings transferred for 3d to high NaCl Adding calcium (Ca2+) to root growth medium enhances salt tolerance in glycophytes (6-8). Ca2+ sustains K+ transport and K+-Na+selectivity in Na+-challenged plants (8) Science 280, 1943-1945
Dealing with Ion Toxicity • Because Na+and K+have similar physic-chemical properties, high concentration of Na+ inhibits K+ uptake by the root. K+ uptake via Arabidopsis KUP1 is inhibited by >5 mM NaCl.Plants use both low and high affinity systems for K+ uptake (to match different soils). • Sodium, once enters into the cytoplasm, inhibits many enzymes. This inhibition is also dependent on the K+ level in the cytoplasm • Na+ are more damaging on the low affinity system that has low K+/Na+ selectivity. Under Na+ stress, it is necessary to use more selective high affinity K+ uptake system in order to maintain adequate . • It is a general phenomenon that salt treatment of plants causes a decrease in cellular K+ content, which may be partly responsible for reduced growth and vigor under salt stress.
High-Affinity Potassium Transporter • AtKUP1 and AtKUP2 Complement Potassium Transport Deficiency in E. coli TK2463 Cells. Enhanced 86Rb+ Uptake in Transgenic Arabidopsis Suspension Cells Expressing AtKUP1
Na+ UPTAKE/EXTRUSION IN THE PLANT CELL Plasma Membrane Na+ Na+ PPi H+ K+ H+ High-affinity K+ transporters V-PPase H+ Na+ Na+/H+ antiport Vacuole Na+ Na+ Tonoplast V-ATPase K+ K+/Na+ selectiveVICs H+ ATP ATP K+/Na+ ratio H+ P-ATPase Adapted from Mansour et al. 2003
The plasma membrane proton H+ pump Plants actively extract nutrients (NPK, etc) from the soil, and actively transport products of photosynthesis (such as sucrose) to parts of the plant that do not carry out photosynthesis (roots). The key enzyme in these processes is the plasma membrane H+-ATPase that pumps protons across the PM and thereby generates the proton and electrical gradient that is the driving force for secondary active transport executed by carriers and channels
Mechanisms of Salt Entry into Root Cells • Current evidence suggests that Na+ enters root cells through various cation channels that could be voltage-dependent or independent cation channels (VIC). Among them, VIC channels are considered the major route for Na+ entry(Amtmann and Sanders, 1999 • Under normal conditions, the plasma membrane potential (MP) of root cells is -130 mV. A more negative potential would facilitate entry of the positively charged Na+ into cells. MP in plant cells is generated by ATPases, which pump H+ out of the cell creating electrochemical potential which facilitate the uptake of solutes. • Some transporters affect salt sensitivity indirectly by altering MP as a result of regulation of ion flux. For example, in yeast trk mutants are defective in K+ uptake, the PM becomes hyperpolarized and this enhances the uptake of cations and rendered the mutants more sensitive to Na+, Li+, and low pH (Serrano et al., 1999 ). • Membrane hyperpolarization enabled K+ uptake through other transporters.Interestingly, Ca2+ can reverse the salt sensitivity in the pmp3mutant. PMP3 is a small hydrophilic protein predicted in the PM. It is not known how this protein can regulate membrane potential. PMP3 is homologous to the Arabidopsis proteins RCI2A and RCI2B (Medina et al., 2001 ; Nylander et al., 2001 )
Sodium Fluxes through Nonselective Cation Channels in the Plasma Membrane of protoplasts from Arabidopsis Roots How do you distinguish Na+ influx catalyzed by NSCCs from that catalyzed by K+-selective and Ca2+-selective channels? Instantaneous currents through the plasma membrane of Arabidopsis root protoplasts in response to voltage-clamp steps from 160 to 80 mV (holding potential = 70 mV). Solutions contained 10, 20, or 100 mM NaCl. To distinguish Na+ influx catalyzed by NSCCs from that catalyzed by K+-selective and Ca2+-selective channels, experiments with K+ and Ca2+ channel blockers Plant Physiol. 2002 February; 128(2): 379–387.
Ion Homeostasis Transport Determinants Plasma membrane: Influx - Na+ influx is passive(nonselective cation channel(s) (NSCC), HKT1 transport system, leak through K+ uptake systems;Cl- uptake is active (because of the inside negative potential across the plasma membrane) Efflux – Na+ efflux is active, H+ driven Na+ antiporter SOS1, proton gradient is established by the plasma membrane (P-type) H+-ATPase, note the ∆pH Tonoplast: transport into the vacuole Na+- influx, H+ driven Na+ antiporter NHX family, proton gradient is established by the tonoplast (V-type) H+-ATPase and pyrophosphatase, ∆pH
perox ATP Ca2+ Osmotic Adjustment and Ion Comparmentalization Cells expend ~50% of their total energy to maintain gradients of ions across membranes. The electrochemical potential of these ion gradients represents stored energy. Plants and fungi are similar in that they use proton (H+) gradients as the "currency" with which to mediate transport of ions K+(Na+) K+(Na+) H+ K+ Na+/H+ K+ polyols proline betaine trehalose pH 7.5 cp Na+ *-scavenging pH 5.5 Tonoplast mt NaCl↑ Plasma Membrane -120 to -200 mV ATP Na+ Cl- Ca2+ H+ ATP pH 5.5 H+ PPi +20 to +50 mV Na+ H+ H+ Ca2+ Na+ H+ H+ H+ H2O Cl- Cl- ATP Cl- Ca2+ H+ Cl- Ca2+ Ca2+ Ca2+ Inositol H2O Na+ Cell volume increases10- to 100-fold during growth and development due almost entirely to an increase in the vacuole size, i.e., water uptake into the vacuole drives cell expansion Na+ and Cl- compartmentalizationin the vacuole is a necessary component of osmotic adjustment, net uptake of these ions across the plasma membrane is restricted and organic osmolytes mediate osmotic adjustment in the cytosol
Model of H+ pumps and transporters found in the plant vacuolar membrane A) H+-ATPase (1) and H+-PPase (2) transport protons (H+) into the vacuolar lumen. Organic and inorganic anions (A) enter the vacuole via channels (4) to electroneutralize, allowing the generation of a pH gradient. This (proton electrochemical gradient [PEG] drives secondary active accumulation of organic and inorganic cations into the vacuole via H+/cation antiporters (3), with osmotically accompanying water. B, Ectopic expression of cation/H+ antiporters (3) in the vacuolar membrane sequester higher amounts of cations through the utilization of the existing PEG generated via the two H+ pumps (1 and 2). C, Reduced H+-pumping activity in the det3 mutant. H+-ATPase activity (1) in the det3 mutants is diminished. A reduction in the PEG activities (3 and 4) across the vacuolar membrane. D, Ectopic expression of AVP1. Transgenic plants with enhanced AVP1(2) have an enhanced PEG. This altered PEG increases transport activities (3 and 4) across the vacuolar membrane.
Transgenic tomato with vacuolar Na+/H+ antiport (AtNHX1) allowed them to grow in 200 mM NaCl vNa/H wt
Ca2+ in Na+ stress • An important determinant for salt tolerance relevant to Na+ and K+ homeostasis is Ca2+. Increased Ca2+ supply has a protective effect on plants under Na+ stress. Early experiments did not distinguish whether Ca2+ acted extracellularlly or intracellularly. Recently, altered cellular Ca2+ homeostasis showed that internal/cytosolic Ca2+ is important to salt sensitivity regulation. 1) e.g., expression of AtACA4 that codes for a vac- Ca2+-ATPase in yeast increased their salt tolerance 2) Arabidopsis vacuolar Ca2+/H+ antiporter gene CAX1, when overexpressed, increased sensitivity to ionic stress. These transgenic plants appeared Ca2+-deficient despite a higher total Ca2+ content(Hirschi,1999) hkt1 mutation suppressed Na+ sensitivity of sos3 mutants, but not in low Ca2+ (0.15 mM), suggesting an alternative Na+-influx system, different from AtHKT1, that is hampered by high Ca2+ (2 mM) but is the prevalent Na+ entry pathway at low external Ca2+.
CAX1 Expression Disturbs Normal Vigor this study shows perturbed growth by constitutive expression of a single transport protein. The CAX1-transgenics displayed altered phenotypes and increased stress sensitivity Plant Cell, Vol. 11, 2113-2122 • CAX1-expressing lines • CAX1-expressing lines after several weeks in the greenhouse. • & (D) Size of CAX1-expressing plants. in the background is expressing CAX1 in the antisense confirmation. This plant is the same size as control plants • (E) The sense roots are significantly stunted. • (F) Leaf of 10-week-old vector control plant grown for 2 weeks without Ca2+. • (G) Leaf of 10-week-old CAX1-expressing plant given Ca2+ supplementation.
Plant phenotypes of altered expression of H+ pumps and H+/cation antiporters Expression of CAX1 in tobacco causes apical burning and other growth defects associated with calcium deficiencies. B, CAX2 makes plants more tolerant of Mn. The CAX2 sense- and antisense- plants grown in MnCl2. Control (C) and AtNHX1 transgenic tomato D) growing in the presence of 200 mM NaCl. E, det3 and control Arabidopsis plants grown in soil. F, Control and transgenic AVP1 lines after recovery from 10 d of drought stress. Gaxiola, R. A., et al. Plant Physiol. 2002;19:967-973
Ion Sensitivity of CAX1-Expressing Plants Two vector control plants are shown at left and two CAX1-expressing plants (35S::CAX1) at right. (A) Plants grown in standard media immediately after transfer to various media (pretreatment). (B) Plants transferred to standard media and grown for 10 days. (C) Plants transferred to standard media supplemented with 50 mM MgCl2 and grown for 10 days. (D) Plants transferred to standard media supplemented with 100 mM KCl and grown for 10 days. (E) Plants transferred to standard media supplemented with 50 mM NaCl and grown for 10 days. (F) Plants transferred to standard media supplemented with 100 mM CaCl2 and grown for 10 days. (G) Plants transferred to standard media without Ca2+ and grown for 10 days. (H)Plantstransferredto standard media supplementedwith 50 mM MgCl2 and 2 mM CaCl2 and grown for 10 days. (I)Plants transferred to standard media supplemented with 100 mM KCl and 2 mM CaCl2 and grown for 10 days
Mol. Cell. Biol. 17, 1289-1297 Salt stress in yeast: the HOG pathway (High Osmolarity Glycerol) of S.cerevisiae The yeast HOG1 signal transduction pathway contains two independent osmosensors. The first is a two-component signal transducer, whereas the second osmosensor, Sho1p, is a transmembrane protein with a cytoplasmic SH3 domain. Under normal osmotic conditions the transmembrane his-kinase Sln1p transfers a phosphate to Ssk1. Phosphorylation of Ssk1p inhibits Ssk1p-mediated activation of Ssk2p and Ssk22p MAPKKKs. Increased osmolarity inactivates Sln1p his-kinase and unphosphorylates Ssk1p activating the Ssk2p and Ssk22p MAPKKKs, which in turn activate Pbs2p. High osmolarity causes Sho1p interaction with and activation of Pbs2p. The activated Pbs2p phosphorylates and activates Hog1p Activation of Hog1p leads to induction of genes for adaptation to high-osmolarity stress, including GPD1, CTT1 and HSP12 primary sensors of osmotic stress, the Sln1p-Ssk1p two-component proteins, are involved in sensing oxidative stress specifically induced by hydrogen peroxide and diamide, but not by other oxidants the HOG1 pathways for adaptation to hyperosmotic stress and the calneurin pathway for ionic stress. In yeast, Na +, K+ and Ca2+ and the pheromone response are regulated by calcineurine; mutants at the calcineurin locus are sensitive to Na+ and Li+.
Membrane stretching in salt stress • In yeast, hyperosmolarity can be sensed by a two-component system composed of the SLN1 His kinase, the YPD1 phosphorelay intermediate, and the SSK1 response regulator, leading to the activation of the HOG1 MAPK pathway. The Arabidopsis (Arabidopsis thaliana) SLN1 homolog, AtHK1, is able to suppress the salt-sensitive phenotype of the yeast double-mutant sln1 sho1 , which lacks both yeast osmosensors (Urao et al., 1999). However, direct evidence for a role of AtHK1 as an osmosensor in plants is still lacking. Although it could interact with the phosphorelay AtHP1 in the yeast two-hybrid system, no interaction was observed between AtHP1 and the response regulators (Urao et al., 2000). Another His kinase, CRE1, which was identified as a cytokinin receptor, is also able to complement the yeast sln1 mutant in the presence of cytokinin (Inoue et al., 2001). Interestingly, a recent work reported that SLN1 and CRE1 perceive the osmotic signal by turgor sensing in yeast (Reiser et al., 2003). It was shown that the integrity of the periplasmic region of SLN1 is essential for its sensor function. This suggests that osmotic stress may trigger a conformational change of SLN1 due to a stress-induced modification of the cell wall-plasma membrane interaction. It is tempting to speculate that a similar turgor-sensing mechanism might regulate hyperosmotic signaling in plants. On the other hand, the involvement of receptor-like kinases (RLK) in osmosensing has been suggested by the increased osmotic stress tolerance induced by overexpression of the tobacco (Nicotiana tabacum) NtC7 (Tamura et al., 2003). • At least two of the Arabidopsis histidine kinase genes, ARABIDOPSIS THALIANA HISTIDINE KINASE1 (ATHK1) and CYTOKININ RESPONSE1 (CRE1), complement sln1 deletion mutants of yeast [15,17] and CRE1 can also respondto changes in turgor pressure when expressed in yeast [15]. Yet, these proteins have not been shown to function asosmosensors in plants
Ion homeostasis after salt (NaCl) adaptation. HOG1 pathways for osmotic homeostasis for (i) low osmolarity sensor SHO1 or (ii) high osmo sensor. SLN1: SLN1 SSK1 PBS2HOG1 or SHO1 PBS2HOG1. Stress adaptation effectorsare those that mediate ion homeostasis, osmolytebiosynthesis, toxic radical scavenging, water transport. Both pathways converge at PBS2 leading to transcriptional activation of glycerol biosynthetic genes High NaCl causes cytosolicaccumulation of Ca2+ and this signals stress responsesthat are either adaptive or pathological. NaCl Ca2+ CDPKs/MAPKs
Activation of two MAPK cascades in yeast HOG MAP kinase cascade Science 299:1025-7 (Left) The mating cascade is activated when the cell's a-factor receptor receives the a-factor pheromone from an expectant partner. The receptor is associated with a G protein, and interaction with pheromone frees the G protein. that exposes a surface which binds to the scaffold Ste5. (Right) High osmolarity cascade is activated by the membrane protein Sho1. Under high-salt conditions, Sho1 exposes a surface that binds to the scaffold Pbs2. (Center) Ste20 is an active kinase tethered to the membrane. Prot-G recruits Ste5 to the membrane, where Ste20 triggers the mating cascade. Sho1 recruits Pbs2 to the membrane, where Ste20 triggers the osmolarity cascade. The Pbs2 scaffold has two bound kinases and akinase domain. Fus3 and Hog1 are called MAPKs, Ste7 and Pbs2 are MAPKKs, and Ste11 is a MAPKKK.
SH3 domains(AtSH3Ps Partially Complement a Salt-Sensitive, Endocytosis-Deficient Yeast Mutant) The basic fold of SH3 domains contains five anti-parallel beta-strands packed to form two perpendicular beta-sheets. The ligand-binding site consists of a hydrophobic patch that contains a cluster of conserved aromatic residues and is surrounded by two charged and variable loops Domain Binding and Function Src-homology 3 (SH3) domains generally bind to Pro-rich peptides that form a left-handed polyPro type II helix, with the minimal consensus Pro-X-X-Pro. Each Pro is usually preceded by an aliphatic residue. Each of these aliphatic-Pro pairs binds to a hydrophobic pocket on the SH3 domain. Class I and 2 of SH3 domains have been defined which recognize RKXXPXXP and PXXPXR motifs respecitvely
Osmotic stress-activated protein kinases in plants MAPKs are induced by osmotic stress (salt and drought) stress FEBS 498;172 (2001) Two MAPKs are activated in an in-gel assay. One is activated at moderate concentrations, responding in a dose-dependent way, peaking at 500 mM NaCl, whereas the other was only activated at very high concentration, starting at 500 mM NaCl The fact that different salt ranges activate different pathways supports the concept that stress is detected by different receptors responding over those limited ranges, in a manner similar to the osmo-sensors in yeast
Salt signaling in yeast & plants In addition to MAPK pathway, yeast has another pathway specific for high NaCl, which includes calcineurin, a phosphatase dependent on Ca2+, and calmodulin. Therefore, it is possible that external high NaCl increases intracellular Ca2+, which then causes calmodulin to transmit signals to other, downstream components, such as calcineurin The SLN1 branch of the HOG pathway is stimulated by turgor reduction An ArabidopsisGSK3/shaggy-Like Gene that Complements Yeast Salt Stress-Sensitive Mutants Is Induced by NaCl and Abscisic AcidPlant Physiol.119 :1527 encode kinases Sln1 and Sho1 have distinct cellular distributions. (A) Architecture of the SLN1 and SHO1 branches of the HOG pathway. (B) Either the SLN1 or SHO1 branch is sufficient to survive on high osmolarity. SLN1 and SHO1 branches in the HOG pathway respond independently to osmotic status of the environment and are apparently redundant. However,in the SLN1 branch, a transmembrane (TM) histidine kinase Sln1 serves as an osmosensor, and transmits the signal through the Sln1–Ypd1–Ssk1 multistep phosphorelay to the redundant pair of kinases Ssk2 and Ssk22. In contrast, another TM protein (Sho1) serves as a facilitator of signaling module assembly that includes Pbs2, Ste11, Ste20, and Cdc42
Activation of distinct lipid and MAPK signalling pathways by osmotic stress Activation of different receptors, dependent on the stress level when 100 mM NaCl is then stressed by additional salt,the same signaling pathways were still activatedin the same response pattern. This is unlikely, if they detect salt concentrations thus, they detect a consequence of increased salt, such as loss of turgor. Thus, osmo-sensors are stretch receptors that respond to changes in membrane pressure More on MAPK MAPKK in salt stress in MAPKs-Plants Sig slide 13 CDPK, calmodulin-like domain protein kinase; DAG, diacylglycerol; DGPP, diacylglycerol pyrophosphate; IP3, inositol 1,4,5-trisphosphate; L-PA, lyso-PA; MAPK, mitogen-activated protein kinase; PA, phosphatidic acid; PI3K, phosphoinositide 3-kinase; PI(3,5)P2, phosphatidylinositol 3,5-bisphosphate; PLA2, phospholipase A2; PLC, phospholipase C; PLD, phospholipase D.
Comparison of Salt Sensitivity in sos mutants on Vertical Plates by Using the Root-Bending Assay Five-day-old seedlings were transferred from normal MS medium to high NaCl, and the seedlings (with roots upside down) were grown for 7 d. Continued growth on salt plates results in bending of roots due to gravitropism; thus,lack of root bending is a visual sign of inhibition by NaCI Genetic analysis indicates that SOS1 is epistatic to SOS2 and SOS3 Zhu Plant Phys 2000
Sensitivity of salt sensitive mutants to other salts sos3 but not sos1 is fixed by high Ca WT, sos1, and sos3 mutants were exposed to high-salt (100 mM NaCl) or low-K+ (20 µM K+) stress
Complementation of sos3 by the wild-type SOS3 : SOS3 Binds 45Ca2+. SOS3 encodes an EF hand–type Ca2+ binding protein with an N-myristoylation domain Similar to B-subunit of calcineurin (type 2B) protein phosphatase G2A (N-myristoylation ) Mutation Abolishes SOS3 Function in Plant Salt Tolerance but not Ca2+ binding SOS3 is a calcium binding protein with an N-myristoylation signature sequence Plant Cell, Vol. 12, 1667-1678
sos1 Plants Cannot Grow with Low K+ suggests the mutant may be deficient in high-affinity K + uptake sos1 needs high levels of K+to grow suggesting that the mutant may be deficient in high-affinity K + uptake The K +content in the wild type did not decrease to <3%of the dry weight,while in sos1 it de-creased to ~1%, indicating that K +deficiency occurs in NaCI-treated sos1 plants. (A)Plants on 20 mM K +. (B)Plants on 200 nM K*.